Биологические основы регенеративной медицины
ВВЕДЕНИЕ
Окружающие нас предметы нередко портятся и ломаются, а также изнашиваются при интенсивном или длительном использовании, поэтому их приходится ремонтировать, заменяя или починяя испорченные части. Иногда это удается и тогда после починки вещь может прослужить еще какое-то время. В отличие от неодушевленных предметов, которые приходится время от времени реставрировать, живые организмы обладают в той или иной степени способностью к самовосстановлению. Феномен самовосстановления живых организмов называют регенерацией (от латинского regeneration, что означает возрождение, восстановление). В биологии термином регенерация называют процессы восстановления организмом утраченных частей тела и органов, а также менее заметные, но не менее важные процессы обновления тканей и клеток. Регенерация является гомеостатическим механизмом, путем, которого организм поддерживает или восстанавливает структуру тканей с тем, чтобы поддержать или восстановить функцию, которую они выполняют (Morgan, 1901 ) Успехи достигнутые в молекулярной биологии, генетике, гистологии, биологии развития, а главное – открытие методов выделения и культивирования стволовых клеток (от нем. Stammzelle – термин введенный Александром Максимовым в 1909 году для клеток которые, делясь обеспечивают постоянную замену изношенных клеток новыми) привели к созданию в 21 веке нового направления медицинской науки: «Регенеративной биологии и медицины» (Stocum, 2006 , Ramires, 2010 , Vogel, 2007 ). Предполагается, что «Регенеративная биология и медицина» может стать важнейшим направлением развития медицины будущего.
Почему это направление столь актуально? Причина кроется в тех медицинских проблемах, которые ученые надеются разрешить с помощью методов регенеративной медицины:
1.) Снабдить организм стволовыми клетками, выращенными вне организма, чтобы помочь ему и облегчить обновление клеток в органах и тканях нуждающихся в ремонте;
2.) Суметь задействовать скрытые регенеративные способности человеческого организма путем направленного воздействия на генетический аппарат, в частности: заставить организм вместо того чтобы идти по пути формирования рубцовой ткани осуществлять полноценную регенерацию с полным восстановлением функций на месте повреждения;
3.) Разработать методы выращивания органов и тканей для трансплантации и таким образом избавиться от проблем поиска доноров и реакций отторжения чужеродного органа;
4.) Найти способы борьбы с функциональными расстройствами, вызванными старческим дряхлением;
5.) Найти способы борьбы с раковыми заболеваниями в частности, найти способы предотвратить переход (трансдифференциацию) эпителиальных раковых стволовых клеток в мезенхимальные, что значительно повышает их инвазивность, способность образовывать метастазы и устойчивость к лекарствам.
Решение столь грандиозных проблем требует глубокого знания фундаментальных процессов лежащих в основе регенерации. Поэтому целью регенеративной биологии и медицины как науки является создание единой фундаментальной картины механизмов регенерации на основе достижений современной биологической науки и использование этих знаний для решения медицинских проблем.
Тот факт, что регенеративная медицина бурно развивается и очевидно в ближайшие 20-30 лет станет одним из ключевых направлений медицины, диктует острую необходимость ознакомить новое поколение медиков со специфической терминологией и основной проблематикой этой науки. Предполагается, что читатель знаком, по крайней мере, со школьным вводным курсом биологии. Цель этой книги будет достигнута, если у читателя возникнет интерес к великим тайнам, сопровождающим деятельность живых организмов и желание заняться проблемами изучения и использования удивительных процессов, составляющих суть сложнейших преобразований происходящих в многоклеточных организмах при регенерации.
Часть I.ОРГАНИЗМЫ – МОДЕЛИ ДЛЯ ИЗУЧЕНИЯ РЕГЕНЕРАЦИИ
правитьГлава1. РЕГЕНЕРАЦИЯ У НАИБОЛЕЕ ПРОСТО УСТРОЕННЫХ ОРГАНИЗМОВ
правитьСпособность различных организмов к регенерации не одинакова. Наиболее просто устроенные организмы имеют, как правило, большие способности к регенерации, чем более сложно устроенные. Поэтому целесообразно начать знакомство с регенерацией на примере более простых организмов, что облегчит понимание этих процессов у более сложно устроенных организмов и в частности у млекопитающих и человека. Приведенные ниже в качестве примеров организмы достаточно легко размножать в условиях самой простой институтской, школьной и даже домашней лаборатории, поэтому они удобны для экспериментального изучения процессов происходящих при регенерации.
Доказательство ведущей роли генетической регуляции в процессах морфогенеза при регенерации на примере одноклеточных. Понятие цитоплазматической сегрегации. Полярность цитоплазмы
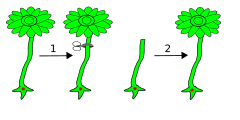
правитьЗамечательную способность организма – замещать утраченные части тела вновь образованными отмечал еще Ч. Дарвин . Он приписывал это свойство действию специфического фактора, присущего всем живым существам, который он называл координирующей силой организма «постоянно стремящейся вновь приводить все части ко взаимной гармонии» Сегодня мы уже знаем, что большинство клеток организма содержит всю необходимую информацию для построения организма заново. Большая часть этой информации заключена в генетическом материале хромосом. Так, например: необходимым условием регенерации амебы является наличие генетического материала в оставшейся части ее тела, которая может составлять лишь 1/80 ее первоначального объема (Vorontsova, Liozner, 1960). Инфузория трубач способна регенерировать из 1/64 ее исходного размера при условии, что этот фрагмент сохранил хотя бы одно звено макронуклеуса (аналог ядра), некоторое количество внутриклеточного материала и часть клеточной оболочки с ресничками (Tartar,1961). В опытах Г. Хеммерлинга (Hammerling 1953) на одноклеточной зеленой водоросли Acetabularia была доказана важная роль нуклеиновых кислот в процессах морфогенеза при регенерации. Хотя весь организм этой водоросли состоит из одной клетки и имеет лишь одно ядро, ацетабулярия достигает нескольких сантиметров в высоту и состоит из трех морфологически хорошо отличимых друг от друга частей: похожей на корень ризоидальной части, которой она прикрепляется к придонным камням, тонкого растущего вверх «стебля» и, похожей на шляпку структуры, расположенной на его вершине. Цитоплазма всех трех частей водоросли не прерывается (см. рис.1а ).
Известно несколько видов ацетабулярии, различающихся по строению шляпки, что позволяет легко их идентифицировать. Если у ацетабулярии A. Mediterranea удалить шляпку, а затем «стебель» водоросли отсечь от своего ризоида содержащего ядро и пересадить на ризоид A. Wettstein (старое название) содержащий «чужое» ядро, то примерно через месяц шляпка регенерирует. При этом вновь образованная шляпка будет иметь черты сходства с обеими родительскими формами. Однако если эту регенерировавшую шляпку удалить, все последующие регенераты внешне ничем не отличаются от того вида, которому принадлежало пересаженное ядро т.е. от A. Wettstein.
Таким образом, информация по строению регенерирующей шляпки находится в ядре и передается от ядра через цитоплазму к вершине стебля, где осуществляется регенерация. Образование первоначально шляпки промежуточного типа являлось, очевидно, следствием стабильности рибонуклеопротеидов (РНП) от старого ядра, в результате чего реализовалась комбинированная инструкция: запасенная в виде РНП информация от старого ядра и вновь образованная информация от нового ядра. При подавлении синтеза РНК актиномицином Д (антибиотик подавляющий синтез нуклеиновых кислот) и обработке стебля ферментом рибонуклеазой, расщепляющей молекулы РНК, формирование шляпки не происходит. Понятно, что наряду с выводом о необходимости синтеза РНК для осуществления процесса формообразования при регенерации шляпки ацетобулярии, возникает и закономерный вопрос о том каким образом синтезированная в ядре РНК приводит к образованию именно шляпки, а не дополнительных ризоидов или стебля. Откуда клетка узнает о том, что повреждена именно шляпка и именно ее необходимо реставрировать. Четкого ответа на этот вопрос пока, к сожалению нет.


Рис 1. а) Ацетабулярии как A. Mediterranea, так и A. Wettstein, представляют собой одноклеточный организм в виде гигантской одноядерной клетки состоящей из ризоида с ядром, стебелька и шляпки; б) Если ацетабулярию A. Mediterranea отсечь от своего ризоида содержащего ядро и пересадить на ризоид A. Wettstein (старое название) содержащий «чужое» ядро, а затем отсечь и шляпку, то примерно через месяц шляпка регенерирует. При этом вновь образованная шляпка будет иметь черты сходства с тем видом, которому принадлежало пересаженное ядро т.е. с A. Wettstein.
Интересно отметить, что регенерация шляпки у ацетобулярии происходит и в том случае если через некоторое время после удаления шляпки перерезать стебель и удалить ризоидальную часть растения. Таким образом, сиюминутного присутствия ядра для регенерации не требуется – достаточно той информации, которую ядро передает стеблю в промежуток между удалением шляпки и удалением ризоидов вместе с ядром. Таким образом, РНК необходимая для регенерации, запасенная в виде РНП стабильна и имеет достаточно длительный период жизни. Закладка специфической организации шляпки происходит намного раньше той стадии, на которой эта форма реализуется. Следовательно, для формообразования помимо регуляции на уровне транскрипции в ядре требуется еще и регуляция на уровне цитоплазмы. Регенерация шляпки на обрезанном с обеих сторон стебле происходит всегда с той стороны стебля, где раннее находилась шляпка, а не ризоиды, что четко свидетельствует о полярной организации цитоплазмы в стебле ацетобулярии. Здесь мы сталкиваемся с явлениями цитоплазматической сегрегации (от лат. Segregare – различие), то есть с различиями цитоплазмы в разных частях клетки. Цитоплазматическая сегрегация, проявляющаяся в полярности цитоплазмы, связана с направленным транспортом РНП, важную роль в котором выполняет цитоскелет (Vogel et al. 2002 ). Полярная ориентация микротрубочек и микрофиламентов цитоскелета по неизвестному пока механизму обеспечивает направленный транспорт необходимых РНП к месту регенерации. Этот же механизм транспорта задействован, очевидно, и тогда когда, по завершении развития ацетобулярии ее ядро делится примерно на 10 000 вторичных ядер, которые мигрируют вверх по стебельку и в радиальные лучи шляпки, где образуются цисты, которые после отмирания шляпки превращаются в снабженные жгутиками гаметы (Bonotto (1994 )).
-
 Удаление зонтика (шляпки). После этого одноклеточное растение формирует новую шляпку
Удаление зонтика (шляпки). После этого одноклеточное растение формирует новую шляпку -
 Ампутация ножки (ризоида), которая содержит единственное клеточное ядро растения. Образуется новый ризоид, который, однако, не имеет ядра. Клетка может выжить в благоприятных условиях несколько месяцев, но уже не способна к размножению
Ампутация ножки (ризоида), которая содержит единственное клеточное ядро растения. Образуется новый ризоид, который, однако, не имеет ядра. Клетка может выжить в благоприятных условиях несколько месяцев, но уже не способна к размножению -
 Стебелёк отрезан ближе к шляпке. Это приводит к образованию второй шляпки
Стебелёк отрезан ближе к шляпке. Это приводит к образованию второй шляпки -
 Стебелёк отрезан чуть выше ризоида. Это приводит к образованию второго ризоида
Стебелёк отрезан чуть выше ризоида. Это приводит к образованию второго ризоида -
 Удаление ризоида и шляпки (оставлена центральная часть). Происходит их восстановление без ядра, но с соблюдением полярности
Удаление ризоида и шляпки (оставлена центральная часть). Происходит их восстановление без ядра, но с соблюдением полярности -
 Удаление шляпки, затем стебель переворачивают и приживляют ризоид с ядром. Если затем через некоторое время снова удалить ризоид с ядром, то шляпка отрастет не с того конца где она была первоначально, т.е. без соблюдения полярности. Из этого эксперимента следует, что ядро отвечает за "обучение".
Удаление шляпки, затем стебель переворачивают и приживляют ризоид с ядром. Если затем через некоторое время снова удалить ризоид с ядром, то шляпка отрастет не с того конца где она была первоначально, т.е. без соблюдения полярности. Из этого эксперимента следует, что ядро отвечает за "обучение".






Литература
править- Лиознер Л.Д., (1962) Восстановление утраченных органов. М., Изд-во АН СССР
- TartarV,(1961) Biology of Stentor. Pergamon Press N.-Y
- Hammerling J. (1953) In: Intern. Rev. Cytol. Vol 2, 475-498
- Vogel H, Gerd E. Grieninger, Klaus H. Zetsche and Linsdley F. (2002), Differential Messenger RNA Gradients in the Unicellular Alga Acetabularia acetabulum. Role of the Cytoskeleton. Plant Physiol, Vol. 129, pp. 1407-1416
- Bonotto S. (1994) Developmental Biology of Acetabularia. J. Mar. Biol. Ass. UK., 74, 93-106
- Bonotto, S., & Luttke, A. (2019). Cellular biology of Acetabularia. In Vol. 2 (pp. 203-246). De Gruyter. https://doi.org/10.1515/9783110837506-013
- Andresen, I. J., Orr, R. J., Shalchian-Tabrizi, K., & Bråte, J. (2021). Compartmentalization of mRNAs in the giant, unicellular green alga Acetabularia acetabulum. Algal Research, 59, 102440. https://doi.org/10.1016/j.algal.2021.102440
Некоторые соматические клетки многоклеточных организмов содержат достаточно информации для построения целого организма.
правитьКаким образом из одной единственной клеточки под названием зигота образуются миллиарды самых разных по своим функциям клеток, образующих органы и ткани, которые слаженно работают, позволяя жить и развиваться нашему организму?
В начале жизни эмбрион образуется как группа клеток, состоящая из трех слоев: внешнего (эктодерма), среднего (мезодерма) и внутреннего (энтодерма). Эти слои развиваются во все органы и ткани у животного.
В 2008 году японский ученый Йошики Сасай (Yoshiki Sasai) обнаружил, что если позволить плюрипотентным стволовым клеткам слипаться с образованием кластеров клеток, они начинают самоорганизовываться в структуры, так называемые эмбриоидные тела. Подобные явления самоорганизации можно увидеть только в группах, насчитывающих приблизительно от 1000 до 100000 клеток. Клетки «сортируются»: однотипные слипаются, а разнотипные остаются разобщёнными. Позднее возникают центры организации, руководящие морфогенезом путём выделения ростовых факторов (морфогенов) с помощью градиентов, концентрации которых создают так называемые биополя.
Этой удивительной особенностью клеток заключающейся в способности к селективной самосборке, их наделяют молекулы адгезии, расположенные на клеточной поверхности. Одна из таких молекул – кадгерин в присутствии кальция подобно клею связывает между собой «подходящие друг другу» клетки с образованием и поддержанием тканей и органов. Клетки, которые имеют один и тот же тип кадгерина, взаимодействуют друг с другом, но клетки с различными типами кадгеринов остаются раздельными. За последние три десятилетия было выявлено множество типов кадгерина и взаимодействующих с ним белков вроде нектинов в различных эмбриональных и взрослых тканях простых и сложных организмов. Показано, к примеру на мозговых клетках, что синтез этих и многих других белков во время онтогенеза регулируется упорядоченным образом, зависящим от фазы развития и старения [1] .
Понять принципы самосборки позволяет аналогия. Каждый, кому довелось покупать в больших супермаркетах, где продаются десятки тысяч различных товаров, видел как быстро, легко и просто кассирша находит информацию о проданном товаре с помощью устройства считывающего его штрих-код. А теперь вспомним замечательный конструктор Лего. В основе каждого набора множество автоматически связывающихся кирпичиков (Automatic Binding Bricks) LEGO, которые, благодаря хитроумным шипам и пространственно соответствующим им комплементарным полостям, легко сцепляются друг с другом. Из подобных кирпичиков-модулей можно построить самые разные модели домов, автомобилей, роботов и других конструкций.
Примерно те же принципы, только значительно усложненные, использует природа для построения живых многоклеточных организмов. Роль кирпичика-модуля здесь выполняет живая клетка. А чтобы каждая клетка нашла свое место в архитектуре организма, используется химический штрих-код, с помощью которого клетки узнают друг друга и сцепляются. Для записи этого кода природа использует сложные углеводы – гликаны, представляющие собой полимеры, состоящие из моносахаридных звеньев.
Еще в 1907 году Вильсон (Wilson) обнаружил поразительный феномен самосборки губок [2]. Он показал, что если дезинтегрировать губку, продавив ее через очень мелкое сито и суспедировать в морской воде, с тем чтобы разделить ее на отдельные клетки и кластеры клеток, а затем дать этой суспензии некоторое время постоять, начинаются удивительные явления. Клетки начинают активно мигрировать и собираются сначала в виде агрегатов в большую неупорядоченную массу, а затем перегруппировываются в организованный многоклеточный организм. В ходе дальнейших экспериментов Вильсон обнаружил еще одно поразительное явление. Оказалось, что если смешать клеточные суспензии двух губок с разной окраской (например: пурпурную Haliclona oculata с красной Microciona prolifera), агрегация приводит к образованию соответственно двух различных губок – красной и пурпурной. Получить химерную губку, в которой были бы клетки обоих видов, Вильсону не удалось.

Вывод из этого эксперимента был очевиден: клетки узнают и агрегируют, взаимодействуя только с клетками своего вида. Позднее выяснилось, что для этого процесса необходимы ионы Ca++, а также видоспецифичные факторы агрегации. Ученые провели эксперимент подтверждающий, что факторы агрегации губок играют важную роль во взаимном узнавании и агрегации клеток. Они выделили протеогликаны из разных видов губок и присоединили их к разноцветным шарикам, изготовленным из латекса, так чтобы протеогликаны каждого отдельного вида, были соединены только с шариком одного цвета. Смешав шарики и продержав их некоторое время в морской воде содержащей ионы Ca++, ученые наблюдали самосортировку шариков – они слипались в комки одного цвета, подтверждая важную роль факторов агрегации губок в видоспецифичном узнавании и агрегации.
Дальнейшие исследования показали, что избирательность к агрегации губок не только видоспецифична, но и распространяется на разные особи одного вида и даже на разные части одного и того же организма. Попытки трансплантировать, то есть пересадить, ткани одного вида губок другому всегда заканчивались отторжением, что свидетельствовало о существовании механизмов распознания губками «своих» и «не своих» [ ]. Множество гликанов, которые выполняют различные функции, такие как, например, механизмы распознания клетками «своих» и «не своих» [ ] находятся и на поверхности клеток млекопитающих.
Важная роль в механизмах опознания этих гликанов отводится семейству Ig-подобных лектинов Siglecs - рецепторов, расположенных на поверхности клеток, опознающих сиаловые кислоты, с помощью своего аминоконцевого домена. По-разному ацетилированные O-ацетил–сиалогликаны расположенные на поверхности клетки, очевидно, играют такую же роль что и штрих-код товаров в супермаркете. Разработана методика позволившая продемонстрировать различия в составе разных O-ацетил–сиалогликанов в зависимости от того где расположена клетка: в каком органе, ткани, типе клетки [ ]. Считается, что Siglecs способствуют межклеточным взаимодействиям, а также необходимы для функционирования врожденной и адаптивной систем иммунитета [ ] . Показано, что имеется корреляция между наличием определенных полиморфизмов в генах кодирующих Siglecs и заболеваниями человека. Так, например, полиморфизм гена CD33 ассоциируется с болезнью Альцгеймера, а полиморфизм гена SIGLEC8 ассоциируется с бронхиальной астмой, SIGLEC14 с хронической обструктивной болезнью легких, полиморфизм гена CD22 с аутоиммунными заболеваниями [ ]. Каждый Siglec обладает уникальным профилем специфичности в отношении типа сиаловой кислоты, ее связи и состава основной гликановой структуры. Они опознают эндогенные сиалогликаны как молекулярный шаблон "свой", по англ. SAMP (self-associated molecular pattern) – что-то вроде паспорта или штрих кода клетки [ ]. Это своего рода сигнальный язык «биологический штрих-код», позволяющий отличать по поверхностным белкам одни клетки от других, кодировать локализацию клетки и ее принадлежность определенным тканям [ ]. Так, общей особенностью, взятых из разных источников, плюрипотентных стволовых клеток, отличающей их от большинства неплюрипотентных клеток, является особый конкретный характер гликозилирования белков их наружней мембраны. В частности, широко используемые маркеры стволовых клеток - антигены специфичные для стадии эмбриона, такие как SSEA-3, SSEA-4, TRA-l-60, TRA-1-81 являются гликанами клеточной поверхности [ ]. Гликопротеин (сиаломуцин) подокаликсин (podocalyxin) обычно расположен только на недифференцированных клетках человека (ИПСК и ЭСК), но не на поверхности дифференцированных соматических клеток. Именно он является носителем кератансульфатов, распознаваемых как антигены TRA-l-60, TRA-1-81 [ ]. Подобный сигнальный язык «биологический штрих-код» используют и соматические клетки.
С целью классификации путем иммунофенотипирования клеток была предпринята попытка создать так называемую систему кластеров дифференцировки (сокращённо CD от англ. cluster of differentiation) для отнесения клеток к тому или иному типу по представленным на клеточных мембранах молекулам-маркёрам. Хотя наличие одного типа CD обычно не позволяет точно определить популяцию клетки (за исключением нескольких примеров), сочетания маркёров позволяют определить её достаточно чётко. Конечно, «биологический штрих-код», который используют клетки, может быть совсем другим, не тем что используют ученые для иммунофенотипирования. Мы находимся только в самом начале пути по изучению межклеточного сигнального языка. Изучение гликанов участвующих в процессах образования из клеток различных структур, и, в конечном счете, органов только начинается. Использование моноклональных антител показало, что в процессе дифференцировки на поверхности клетки одни гликаны сменяют другие [ ]. Более того соседние клетки могут влиять на метаболизм и судьбу клетки модифицируя рецептор на ее поверхности, например фукозой [ ].
Модификации поверхностных белков, очевидно, участвуют и в отбраковке клеток (в том числе постаревших). Так, анализ сахарных цепей N-связанных с аспарагином, у здоровых старых людей, долгожителей и их потомства, а также мышей, которым ограничили калорийность питания, позволил выявить новых кандидатов в биомаркеры старения, таких как N-гликаны, лишенные остатков галактозы на их ответвлениях [ ]. Эти агалактозилированные биантенальные структуры IgG, названные IgG-G0, в основном декорируют Asn297 в Fc части молекулы. Возрастное накопление IgG-G0 может способствовать хроническому слабо выраженному провоспалительному состоянию, характерному для пожилых людей. Изменения гликозилирования с возрастом, были воспроизведены в широкомасштабных исследованиях населения [ ]. Они позволили выявить строгую зависимость галактозилирования от возраста. Содержание негалактозилированных гликанов (А2 и FA2) постоянно увеличивается с возрастом, в то время как уровень дигалактозилированных гликанов (A2G2, FA2G2, A2BG2, и FA2BG2) с возрастом снижается. Сочетание трех гликанов IgG (FA2B, FA2G2 и FA2BG2) позволяло предсказать хронологический и биологический возраст, значительно точнее других маркеров биологического возраста, таких, как например, длины теломер [ ]. Предполагается, что галактозилирование иммуноглобулинов играет роль защиты («документа с правом на проживание») от активации антитело-зависимой клеточной цитотоксичности.
Одним из маркеров старения является β-галактозидаза – роль которой, очевидно, в изменении «паспортных данных» клетки подлежащей отбраковке, поскольку ее активность отмечается и в зоне висцеральной энтодермы ранних эмбрионов, что никак не связано с хронологическим старением [ ]. Там она очевидно помечает клетки на отбраковку в ходе морфологических изменений, таких как например исчезновение перепонок между пальцами [ ].
Удивительную способность клеток к самосборке используют для выращивания в трёхмерных клеточных культурах (3D cell culture) "затравок" органов названных органоидами. Простейшую форму органоида представляют собой так называемые эмбриоиды (embryoid body). Они используются для первоначальной дифференцировки ИПСК в различные клеточные линии, например в гепатоциты [ ]. Исследование «поведения» органоидов позволит понять сложные молекулярные процессы, благодаря которым происходит построение человеческого эмбриона. В частности, почему вместо хаотического нагромождения, из клеток с той или иной специализацией дифференцировки, образуются хорошо воспроизводимые морфологические структуры соответствующего органоида: либо печени, либо почек, либо сердечно-сосудистой ткани, поджелудочной железы, легочной ткани, тимуса, слезных желез, мозга, сетчатки глаза, и т.д. [ ].
Связующим звеном, обеспечивающим контроль и регуляцию поведения клетки целостным организмом, служит микроокружение клетки - ниша. Ниша обеспечивает клетку факторами, необходимыми для её жизнедеятельности, регулирует ее ориентацию и местоположение в тканевом компартменте. На протяжении развития, клетки воздействуют на поведение друг друга и принимают решения, исходя из "разговора" с соседями и «подсказки» от межклеточного матрикса направляющей дифференцировку клеток в том или ином направлении.
Сократительный актиновый цитоскелет клетки связывается с внеклеточным матриксом с помощью, так называемого фокального контакта, что позволяет клетке «чувствовать» поверхность внеклеточного матрикса и регулировать полярность митоза [ ]. Отделение клетки от взаимодействующего с ней матрикса может привести к гибели клетки в результате аноикиса (англ. anoikis) — запрограммированной смерти клетки в результате утраты нормального взаимодействия с матриксом [ ]. Межклеточный матрикс может воздействовать на клетку выделяя сигнальные молекулы или просто физически – жесткость подложки влияет на судьбу клетки [ ]. Так например Шкуматову с соавт. . удалось получить кардиоваскулярные органоиды, способные поддерживать биения путем культивирования эмбриоидов на коллаген-конъюгированных гидрогелях с жесткостью, подобной жесткости сердечной мышечной ткани [ ]. Очевидно способность клеток реагировать на механические воздействия играет важную роль в морфогенезе и поэтому при культивации клеток вне организма необходимо стремиться к созданию комфортных для клеток механических напряжений, путём регуляции жесткости материала подложек и среды для инкубации [ ]. Так, например, разработан метод, который способствует превращению соматических клеток в стволовые клетки «сжимая» их с помощью 3D микроокружения состоящего из специально подобранного геля [ ].
Хорошо известно, что в развивающихся тканях клетки сравнивают свою пригодность со своими соседями, и менее подходящие («проигравшие») клетки устраняются через явление, известное как конкуренция клеток - механизм контроля качества, который устраняет менее подходящие клетки [ ].
Литература
править- Shiotani, H., Maruo, T., Sakakibara, S., Miyata, M., Mandai, K., Mochizuki, H., & Takai, Y. (2017). Aging‐dependent expression of synapse‐related proteins in the mouse brain. Genes to Cells, 22(5), 472-484. DOI: 10.1111/gtc.12489
- Wilson H. V., (1907). On some phenomena of coalescence and regeneration in sponges. J. exp. Zool., 5: 245-258
- Elda Gaino , Giorgio Bavestrello & Giuseppe Magnino (1999). Self/non-self recognition in sponges, Italian Journal of Zoology, 66:4, 299-315, DOI: 10.1080/11250009909356270
- André, S., Kaltner, H., Manning, J. C., Murphy, P. V., & Gabius, H. J. (2015). Lectins: Getting familiar with translators of the sugar code. Molecules, 20(2), 1788-1823; doi:10.3390/molecules20021788
- Langereis, M. A., Bakkers, M. J., Deng, L., Padler-Karavani, V., Vervoort, S. J., Hulswit, R. J., ... & de Groot, R. J. (2015). Complexity and Diversity of the Mammalian Sialome Revealed by Nidovirus Virolectins. Cell Reports. DOI: http://dx.doi.org/10.1016/j.celrep.2015.05.044
- Bochner, B. S., & Zimmermann, N. (2015). Role of siglecs and related glycan-binding proteins in immune responses and immunoregulation. Journal of Allergy and Clinical Immunology, 135(3), 598-608.
- Schwarz, F., Fong, J. J., & Varki, A. (2015). Human-Specific Evolutionary Changes in the Biology of Siglecs. In Biochemical Roles of Eukaryotic Cell Surface Macromolecules (pp. 1-16). Springer International Publishing.
- Crocker, P. R., Paulson, J. C., & Varki, A. (2007). Siglecs and their roles in the immune system. Nature Reviews Immunology, 7(4), 255-266.
- Angata, T. (2014). Associations of genetic polymorphisms of Siglecs with human diseases. Glycobiology 24 (9): 785-793. doi: 10.1093/glycob/cwu043
- Padler-Karavani, V., Hurtado-Ziola, N., Chang, Y. C., Sonnenburg, J. L., Ronaghy, A., Yu, H., ... & Angata, T. (2014). Rapid evolution of binding specificities and expression patterns of inhibitory CD33-related Siglecs in primates. The FASEB Journal, 28(3), 1280-1293. doi:10.1096/fj.13-241497
- Medzihradszky, K. F., Kaasik, K., & Chalkley, R. J. (2015). Tissue-specific glycosylation at the glycopeptide level. Molecular & Cellular Proteomics, mcp-M115. doi:10.1074/mcp.M115.050393mcp.M115.050393.
- Thomson, J. A., Itskovitz-Eldor, J., Shapiro, S. S., Waknitz, M. A., Swiergiel, J. J., Marshall, V. S., & Jones, J. M. (1998). Embryonic stem cell lines derived from human blastocysts. Science, 282(5391), 1145-1147..
- Toyoda, H., Nagai, Y., Kojima, A., & Kinoshita-Toyoda, A. (2017). Podocalyxin as a major pluripotent marker and novel keratan sulfate proteoglycan in human embryonic and induced pluripotent stem cells. Glycoconjugate Journal, 34, Issue 2, pp 139–145 doi:10.1007/s10719-016-9757-0
- Kandasamy, M., Roll, L., Langenstroth, D., Brüstle, O., & Faissner, A. (2017). Glycoconjugates reveal diversity of human neural stem cells (hNSCs) derived from human induced pluripotent stem cells (hiPSCs). Cell and Tissue Research,368(3), 531-549. doi:10.1007/s00441-017-2594-z
- Kakuda, S., & Haltiwanger, R. S. (2017). Deciphering the Fringe-Mediated Notch Code: Identification of Activating and Inhibiting Sites Allowing Discrimination between Ligands. Developmental Cell, 40 (2), 193–201 https://doi.org/10.1016/j.devcel.2016.12.013
- Parekh R, Roitt I, Isenberg D, Dwek R, Rademacher T (1988). Age-related galactosylation of the N-linked oligosaccharides of human serum IgG. J Exp Med;167:1731–1736. doi:10.1084/jem.167.5.1731
- Vanhooren V, Dewaele S, Libert C, et al. (2010). Serum N-glycan profile shift during human ageing. Exp Gerontol. ;45:738–743. doi:10.1016/j.exger.2010.08.009
- Ruhaak LR, Uh HW, Beekman M, et al. (2011). Plasma protein N-glycan profiles are associated with calendar age, familial longevity and health. J Proteome Res.;10:1667–1674. doi:10.1021/pr1009959
- Kristic, J., Vuckovic, F., Menni, C., et al., & Lauc, G. (2014). Glycans Are a Novel Biomarker of Chronological and Biological Ages. The journals of gerontology. Series A, Biological sciences and medical sciences., 69 (7): 779-789. doi: 10.1093/gerona/glt190
- .Dall’olio F, Vanhooren V, Chen CC, Slagboom PE, Wuhrer M, Franceschi C (2013) N-glycomic biomarkers of biological aging and longevity: a link with inflammaging. Ageing Res Rev. ;12:685–698. doi:10.1016/j.arr.2012.02.002
- Huang, T. and Rivera-Pérez, J. A. (2014), Senescence-associated β-galactosidase activity marks the visceral endoderm of mouse embryos but is not indicative of senescence. Genesis, 52: 300–308. doi: 10.1002/dvg.22761
- Lorda-Diez CI, Garcia-Riart B, Montero JA, Rodriguez-León J, Garcia-Porrero JA, Hurle JM (2015). Apoptosis during embryonic tissue remodeling is accompanied by cell senescence. Aging (Albany NY) 7(11):974–985
- Pettinato, G., Ramanathan, R., Fisher, R. A., Mangino, M. J., Zhang, N., & Wen, X. (2016). Scalable Differentiation of Human iPSCs in a Multicellular Spheroid-based 3D Culture into Hepatocyte-like Cells through Direct Wnt/β-catenin Pathway Inhibition. Scientific Reports, 6, Article number: 32888 doi:10.1038/srep32888
- Dahl-Jensen, S., & Grapin-Botton, A. (2017). The physics of organoids: a biophysical approach to understanding organogenesis. Development, 144(6), 946-951; doi: 10.1242/dev.143693
- Przybyla, L., Lakins, J. N., & Weaver, V. M. (2016). Tissue mechanics orchestrate WNT-dependent human embryonic stem cell differentiation. Cell Stem Cell,19(4), 462-475.
- Ishikawa, F., Ushida, K., Mori, K., & Shibanuma, M. (2015). Loss of anchorage primarily induces non-apoptotic cell death in a human mammary epithelial cell line under atypical focal adhesion kinase signaling. Cell Death & Disease, 6(1), e1619; doi:10.1038/cddis.2014.583
- Hongwei Lv, H., Wang, H., Zhang, Z., Yang, W., Liu, W., Li, Y., & Li, L. (2017). Biomaterial stiffness determines stem cell fate. Life Sciences, 178 , 42-48 DO 10.1016/j.lfs.2017.04.014
- Shkumatov A, Baek K, Kong H (2014) Matrix Rigidity-Modulated Cardiovascular Organoid Formation from Embryoid Bodies. PLoS ONE 9(4): e94764 https://doi.org/10.1371/journal.pone.0094764
- Evans, N. D. and Tusan, C. G. (2017). Extracellular Matrix Structure and Stem Cell Mechanosensing, in Mechanobiology: Exploitation for Medical Benefit (ed S. C. F. Rawlinson), John Wiley & Sons, Inc., Hoboken, NJ, USA. doi: 10.1002/9781118966174.ch1
- Wong, S. Y., Soto, J., & Li, S. (2017). Biophysical regulation of cell reprogramming. Current Opinion in Chemical Engineering, 15, 95-101. https://doi.org/10.1016/j.coche.2017.01.001
- Massimiliano Caiazzo, Yuya Okawa, Adrian Ranga, Alessandra Piersigilli, Yoji Tabata, Matthias P. Lutolf (2016). Defined three-dimensional microenvironments boost induction of pluripotency. Nature Materials 15, 344–352 doi:10.1038/nmat4536
- Merino, M. M., Levayer, R., & Moreno, E. (2016). Survival of the Fittest: Essential Roles of Cell Competition in Development, Aging, and Cancer. Trends in cell biology, 26(10), 776-788.
Понятие плюрипотентности на примере растительных клеток
правитьВ определенных условиях многие из зрелых растительных клеток сохраняют способность делиться и способность менять свою специализацию. Если культивировать растительную ткань в среде, содержащей необходимые питательные вещества и гормоны (в первоначальных экспериментах к культуральной среде добавляли молоко кокосовых орехов) многие из ее клеток начинают бурно делиться и в конце концов образуют массу относительно неспециализированных клеток называемую каллусом (Steward, 1968 ). В зависимости от соотношения фитогормонов культивируемые клетки могут дать начало клеткам корней или побегов. Каллус можно разделить на одиночные клетки и выращивать в суспензионной культуре. Из одиночных клеток, выделенных из суспензии, иногда удается вырастить целое растение. Часто для этого требуется использовать растение-«няньку», которое снабжает росток необходимыми питательными веществами и фитогормонами. Выращивание растений из каллуса или суспендированных клеток сейчас нередко используют для быстрого размножения ценных растений, например женьшеня. Способность одной единственной зрелой соматической (не половой) клетки дать начало целому растению, в котором можно найти все обычные для данного растения типы специализированных (дифференцированных) клеток, показывает, что при специализации (дифференцировке) у этих соматических клеток растения не происходит утраты или необратимой инактивации каких – либо генов. Клетка, способная дать начало целому организму называется плюрипотентной (от лат. pluralis – множественность и от лат. potential – возможность, скрытая способность). Позднее мы увидим, что плюрипотентность свойственна не только культивируемым растительным клеткам, но и некоторым, особым образом обработанным, культивированным соматическим клеткам взрослого организма животных и человека, что имеет очень важное значение для медицины. Особый случай морфогенеза растительной ткани представляют галлы. Галл (от лат. galla — чернильный орешек) — это патологическое образование на органе растения. Их возбудителями могут быть вирусы, бактерии, грибы, но чаще поражения наносятся членистоногими (клещами[1], тлями, галлицами, орехотворками) или корневыми галлообразующими нематодами[2]. При этом поражаются участки листьев и других органов растений. По своей форме и морфологии они резко отличаются от тканей растения. Так, например, галлы, образование которых обусловлено деятельностью паразитирующих на растении насекомых, имеют геометрические формы никак не похожие на нормальные части растения. Галлы это прекрасный образец тканевой инженерии – модификации природных структур, которая возникает благодаря направленному влиянию посторонних организмов. Важным отличием галлов от опухолей вызванных бактериями и вирусами является жесткая регуляция у галлов направленного морфогенеза и процессов дифференциации. Галлы образованные насекомыми относящимися к разным видам могут значительно отличаться по своей морфологии. Так например, личинка . Izeniola obesula внедрившись в паренхиму сердцевины растения вызывает интенсивную клеточную пролиферацию (размножение клеток) и индуцирует дифференцировку клеток в новые сосудистые ткани и оболочку, состоящую из склеренхимы вокруг ее камеры. Личинка Stefaniola defoliate располагается во флоеме и вызывает пролиферацию паренхимы флоемы и лишь позднее окружает себя капсулой из вторичной ксилемы (Dorchin 2002 ). Растение пытается регенерировать повреждения вызванные личинками, а личинка направляет эту регенерацию в своих целях. Есть данные о том, что личинка выделяет определенные морфогенетические факторы, вызывающие изменения дифференцировки клеток растения (Anantanarayanan 2011 ). Предложен гипотетический механизм морфогенеза галла вызванного личинкой. В результате деятельности личинки вокруг нее повышается осмотическое давление. Этот осмотический шок приводит к поступлению воды, которая вызывает расширение полости которую занимает личинка и, как результат регенераторной реакции растения на повреждение к усиленной пролиферации и поляризации клеток окружающих личинку (Anantanarayanan 2011). Вышеизложенное делает каллус и галлы перспективными моделями для изучения морфогенеза при регенерации растений и разработки стратегии тканевой инженерии инкапсуляции для терапии хронических болезней.
Цитоплазматическая сегрегация MDR белков предопределяет продолжительность жизни клетки и ее способность делиться. На примере пекарских дрожжей Saccharomyces cerevisieае
правитьСемейство белков, отвечающих за транспорт молекул через клеточную мембрану, известных под названием MDR ( multidrug resistance proteins ) за их способность помогать раковым клеткам избежать гибели от противораковых лекарств, а в нормальных клетках помогающие избавиться от токсинов, было найдено практически во всех известных организмах от одноклеточных дрожжей до высокоорганизованных организмов включая человека (Ernst et al., 2005 ). Эволюционная консервативность этих белков свидетельствует об их важнейшей роли в жизнедеятельности клеток. Было показано, что при делении дрожжевых клеток происходит неравномерное распределение MDR белков. У материнской клетки остаются все старые MDR белки, тогда как дочерней клетке достаются все новосинтезированные MDR белки. Запас MDR белков у материнской клетки, таким образом, не обновляется – она вынуждена использовать только те MDR белки, которые достались ей при рождении. За время жизни клетки эти белки распадаются и перестают выполнять свою функцию, в результате чего в клетке начинают быстрее накапливаться токсины ограничивающие продолжительность ее жизни и способность делиться ( Eldakak et al., 2010 ). Так, «материнская» клетка дрожжей может дать начало лишь 20-30 дочерним клеткам, прежде чем она теряет способность делиться и погибает (Willyard C. 2010 ). Механизм цитоплазматической сегрегации, посредством, которого происходит неэквивалентное распределение старых и новосинтезированных MDR белков между материнской и дочерней клеткой пока не ясен. Не исключено, однако, что подобный механизм лежит в основе потери старыми многоклеточными организмами, включая человека, способности эффективно обновлять ткани и клетки (Bunting 2002 ), что, в частности, обусловлено потерей стволовыми клетками способности к делению после 30-50 циклов образования дочерних клеток (Budovsky ).
Дифференциация клеток направляется с помощью механизмов цитоплазматической сегрегации на примере нейробластов кузнечика и клеток нематоды C. elegans
правитьВ 1952 году был проведен эксперимент, наглядно демонстрирующий важнейшую роль цитоплазматической сегрегации в механизмах клеточной дифференцировки (специализации). Используя очень тонкую иглу экспериментатор сумел повернуть метафазное веретено делящихся нейробластов таким образом, что хромосомы, которые должны были мигрировать в дочернюю клетку, предназначенную для специализации в ганглий, попали в ту клетку, что была предназначена для образования нейробласта и наоборот, хромосомы, предназначенные для нейробласта, попали в то место где должна была образоваться клетка ганглия. Несмотря на то, что хромосомы оказались не в своей клетке, ничего не изменилось в специализации клеток (Carlson, 1952 ).Таким образом, дифференциация определяется не хромосомами, а той частью цитоплазмы, которая достается при делении дочерним клеткам. Очевидно, что еще до самого деления участок цитоплазмы предназначенный для ганглия получил необходимую информацию-инструкцию для детерминации (от лат.determinare – предопределять) в ганглий, а участок предназначенный для нейробласта инструкцию для образования нейробласта. Следовательно, клетка дифференцируется в соответствии с механизмами цитоплазматической сегрегации. Очевидно, что цитоплазматические факторы предопределяют дальнейшую активность генов хромосомы. Вместе с тем, как мы видели ранее, хранилищем информации необходимой для морфогенеза являются хромосомы. Молекулы, ответственные за детерминацию, переходят при делении только в одну из дочерних клеток, что связано с крайне асимметричным неслучайным распределением цитоплазмы и клеточных органелл при делении клеток (Errington et al., 2010 ). Такие молекулы наделяют клетку способностью к экспрессии (от лат. Expressio - выражать) определенного набора генов необходимого для ее специализации. На примере нематоды C. elegans было показано, что дифференцировка клеток может продолжаться даже после блокады клеточного деления смесью колхицина и цитохолазина В в результате которого разделение цитоплазмы прекращается, но не затрагивается синтез нуклеиновых кислот. При этом у одной клетки появляется целый набор свойств, в норме характерных для разных специализированных потомков этой клетки (Laufer et al.,1980 ).
Литература
править- Ernst R, Klemm R, Schmitt L, Kuchler K. (2005) Yeast ATP-Binding Cassette Transporters: Cellular Cleaning Pumps. Methods in Enzymology.;400:460–484.
- Eldakak A., Rancati G., Rubinstein B., Paul P., Conaway V., Li R., (2010) Asymmetrically inherited multidrug resistance transporters are recessive determinants in cellular replicative aging. Nature Cell Biology. 12, 799–805, doi:10.1038/ncb2085
- Willyard C.(2010) Of cellular dynamics. Deccan Herald content 90834
- Bunting KD. (2002) ABC Transporters as Phenotypic Markers and Functional Regulators of Stem Cells. Stem Cells;20:11–20
- Budovsky A., Vadim E. Fraifeld V.E., Aronov S (2010) Biogerontology. Linking cell polarity, aging and rejuvenation DOI: 10.1007/s10522-010-9305-4
- Carlson J.G. (1952) Microdissection studies of the dividing neuroblast of the grasshopper … .Chortophaga virifasciata. Chromosoma, 5, 199-220.
- Errington R.J. Summers H.D. (2010) Single cell nanoparticle tracking to model cell cycle dynamics and compartment inheritance. Cell Cycle, 9(1), 121-130.
- Laufer J., Barricalupo P., Wood W.B (1980) Segregation of developmental potential in early embryos of Caenorhabditis elegans. Cell , 19, 569-577.
Пути изучения морфогенеза на примере слизистого грибка миксомицета Dictyostelium discoideum
правитьКлючевые слова: морфогенез, регенерация, слизень, smug, миксомицет, Dictyostelium discoideum, центр агрегации, цАМФ, миграции клеток, компенсаторная трансдифференциация, Копин А, Copine A, апоптоз, эпителий, альфа- и бета-катенины, актин.
В генетической программе живых существ записаны присущие им и их потомкам черты, вплоть до крохотного пятнышка на раковине моллюска или папиллярного узора на пальцах человека (который с фантастической точностью восстанавливается после небольших повреждений, свидетельствуя о том, что заданная генетическим кодом, трехмерная структура тканей организма сугубо индивидуальна). Мы легко различаем своих знакомых и родственников по характерным чертам их лица. С помощью генного анализа можно точно установить на уровне ДНК родственные связи. А вот каким «языком» запрограммирована пространственная запись, которая "переводит" химический язык генетического кода в реальную объемную структуру, в тело? Как клетка «узнает», где ей надо остановиться, когда перестать делиться и какую форму принять, чтобы войти в состав сложнейшей архитектуры того или иного органа? Удобным объектом для изучения механизмов морфогенеза (строительства тканей) является слизистый грибок миксомицет Dictyostelium discoideum, генетический код которого примерно в 100 раз проще, чем у человека, размещается только на шести хромосомах и полностью «прочитан» еще в 2005 году. У этого грибка любопытный жизненный цикл, благодаря чему он является излюбленной моделью для изучения процессов морфогенеза и, в частности, процесса образования бластемы (Goss, 1994 ) Сначала его клетки передвигаются в виде "амеб" по почве и делятся через каждые несколько часов. Затем, при истощении запасов пищи, одна или несколько клеток образуют центр агрегации и выделяют цАМФ, что служит сигналом «все ко мне». В ответ на секрецию цАМФ сотни и даже тысячи "амеб" начинают скоординированное движение к центру агрегации, при этом у них еще активируются тысячи новых генов, и они начинают синтезировать вещества необходимые для узнавания и межклеточной адгезии их мембран.
Рис 4. В плазмодии происходят сложные миграции клеток: клетки задней части слизевика образуют подошву для закрепления на почве; клетки передней части слизня перемещаются вниз и образуют ножку, а клетки средней части мигрируют вверх и дифференцируются в споры, образующие плодовое тело
Рис.5 Жизненный цикл слизистого грибка миксомицета Dictyostelium discoideum. Сначала его клетки передвигаются в виде "амеб"по почве. Затем при истощении запасов пищи, одна или несколько клеток образуют центр агрегации и выделяют цАМФ, что служит сигналом «все ко мне». В ответ на секрецию цАМФ тысячи "амеб" начинают скоординированное движение к центру агрегации, слипаются и образуют многоклеточный плазмодий, который выглядит червеобразным слизнем. Этот слизень ползет к свету и затем превращается в маленький тонконогий грибок с круглой головкой, где находятся споры. Из спор прорастают новые амебы.
"Амебы" сползаются, слипаются и образуют многоклеточный плазмодий, который выглядит червеобразным слизнем. Этот слизень выползает на сухое место, ориентируясь на тепло и свет. Интересно отметить, что внешний источник свечения не только служит ориентиром для передвижения, но и задает полярность последующей дифференциации клеток D. Discoideum.
По мере передвижения в нем запускается процесс дифференцировки клеток(Takeuchi et al., 1994 ). В плазмодии происходят сложные миграции клеток: клетки задней части слизевика образуют подошву для закрепления в выбранном месте; клетки передней части слизня перемещаются вниз и образуют ножку, а клетки средней части мигрируют вверх и дифференцируются в споры, образующие плодовое тело (рис 4). Так слизень превращается в маленький тонконогий грибок с круглой головкой, где находятся споры. Прямо-таки на глазах из клеток собирается причудливый организм (рис.5).
Если наполовину сократить количество сливающихся клеток, получается той же формы грибок, только вдвое меньше. Если оставить 1/4 клеток, они, опять, слившись, дают грибок со всеми присущими ему формами, только еще меньших размеров. Как принимается коллективное решение о форме, которую им надо сложить, собравшись вместе? Пропорция между клетками слизня четко регулируется. Удаление какого–либо типа клеток ведет к компенсаторной трансдифференциации. Вместе с тем существуют определенные пределы такой трансдифференциации - «зоны толерантности» к повреждению, разные для разных частей слизня. Так, после ампутации всего заднего участка, трансдифференциация прекращается, если число клеток из которых должен образоваться стебелек сокращается до 10-20% от нормы. Тогда как трансдифференциация при удалении переднего участка (клеток, которые образуют подошву) прекращается, только если число клеток, ниже 10%. При этом морфология слизня регулируется всегда таким образом что соотношение длины к ширине переднего участка постоянно (Rafols et al , 2001 ).
Недавно выяснилось, что важную роль в преобразовании слизня в грибок играет эволюционно консервативный (а значит выполняющий важную функцию как у низших, так и у высших организмов) связанный с клеточной мембраной кальций-зависимый белок Копин А (Copine A). Мутанты не имеющие гена Копина А не способны перейти от стадии слизня к образованию грибка и остаются на стадии слизня. Если их выращивать в условиях дефицита ионов Са, то они образуют грибки с очень коротенькой ножкой. Очевидно, морфогенез стебелька регулируется каким-то кальций зависимым негативным механизмом (Smith T.H. 2010 )
Интересно отметить, что для правильного расположения и формирования комплексной структуры грибка D. Discoideum необходим апоптоз. Около 20% клеток в D. discoideum альтруистично самоуничтожаются при формировании зрелого плодового тела. Клетки, образующие ножку формируют вакуоли и удлиняются, поднимая плодовое тело. По достижении определенной высоты предопределенной «планом» они претерпевают апоптоз и отмирают. Клетки плодового тела, превратившись в споры, рассеиваются ветром и становятся новыми «амебами» (рис.5). .
Механическую прочность высокой и тонкой ножке придает центральная (осевая) трубка, содержащая целлюлозу и некоторые белки. Эти вещества выделяются клетками, образующими нечто похожее на окружающий трубку однослойный эпителий - один из основных типов тканей, характерных для многоклеточных животных (Metazoa). Эпителий состоит из слоя прочно сшитых друг с другом клеток, способных к направленному поглощению и выделению различных веществ. Главной особенностью этих эпителиальных клеток является их поляризованность - расположение внутриклеточных структур и мембранных белков в них строго упорядоченно - центросома и аппарат Гольджи в них всегда располагаются между ядром и поверхностью, обращенной к осевой трубке, на этой же поверхности расположены молекулы фермента, синтезирующего целлюлозу (cellulose synthase). Ключевую роль в формировании этого эпителия играют альфа- и бета-катенины (β-Catenin), белки которые, как известно, участвуют в агрегации и поляризации эпителиальных клеток и у животных. Известно,что альфа-катенины (Alpha-catenin), у животных связываются с актином - одним из основных двигательных белков цитоскелета. Гомолог альфа-катенина.найден и у D. Discoideum. Выяснилось, что этот белок производится в малых количествах, пока амёбы живут поодиночке, но его синтез резко усиливается при формировании многоклеточных структур. Как и альфа-катенин животных, этот белок способен прикрепляться, во-первых, к волокнам актина, во-вторых - к бета-катенину (в опытах использовался, с одинаковым успехом, бета-катенин диктиостелиума и мыши). В экспериментах с поочередным отключением генов альфа- и бета-катенина было показано что оба эти белка необходимы для нормального развития эпителия. Без них клетки не могут должным образом поляризоваться и самоорганизоваться в эпителий, а без эпителия не формируется осевая трубка. В итоге развитие плодового тела обрывается на ранней стадии (Dickinson et al 2011 ).
Литература
править- . Goss R.J.(1994) Developmental decisions in D. Discoideum. Ann Rev. Physiology. 58, 330-357.
- . Takeuchi I., Tasaka M., Okomoto K.,Maeda Y.(1994) Regulation of cell differentiation and pattern formation in Dictyostelium development. Int J.Dev.Biol ;38(2):311-319
- . Rafols I. Amagi A. Maeda Y., MacWilliams H.K., Sawada Y. (2001) Cell type proportioning in Dictyostelium slugs: lack of regulation within a 2.5-fold tolerance range. Differentiation 2001 Jun;67(4-5):107-116
- . Smith T.S., Pineda J.M., Donaghy A.C., Damer C.K. (2010) Copine A plays a role in the differentiation of stalk cells and the initiation of culmination in Dictyostelium development. BMC Dev Biol. 2010; 10: 59.
- . Daniel J. Dickinson, W. James Nelson, William I. Weis.(2011) A Polarized Epithelium Organized by а- and β -Catenin Predates Cadherin and Metazoan Origins. Science. 2011. V. 331. P. 1336-1339.
Исследования генетических механизмов регенерации с помощью интерактивных РНК на примере нематоды Caenorhabditis elegans
правитьCaenorhabditis elegans крошечный червь, длина которого достигает 1 мм. Он имеет сравнительно несложное анатомическое строение (рис 10). Несмотря на то, что он состоит приблизительно всего лишь из одной тысячи соматических и примерно двух тысяч половых клеток, общий план строения его тела в основных чертах тот же что и у большинства высших животных. Тело его имеет удлиненную форму с билатеральной (двусторонней) симметрией. С апикальной (передней) стороны находится глотка, через которую в кишечник засасываются бактерии, а поблизости от заднего конца – анус для удаления испражнений. Снаружи тело покрыто «кожей» - гиподермой, которую подстилает мышечный слой. Помимо кишечника внутри тела имеется трубчатая гонада, построенная из соматических клеток. Внутри гонады расположены половые клетки. Тело червя прозрачно, что позволяет прижизненно наблюдать деление, миграцию и дифференцировку клеток. Важной особенностью C. Elegans является чрезвычайная точность и предопределенность всех процессов развития и регенерации, что позволяет с большой точностью предсказать судьбу каждой отдельной клетки. C. elegans имеет одну из самых «простых» нервных систем (простыми часто называют нервные системы, состоящие из небольшого числа нейронов). Взрослая гермафродитная особь имеет всего 302 нейрона, связи между которыми были полностью описаны. (White et al, 1986 )
Рис.6. Общий план строения Caenorhabditis elegans в основных чертах тот же что и у большинства высших животных.
Генетический аппарат Caenorhabditis elegans расположен на шести парах гомологичных хромосом. В гаплоидном геноме Caenorhabditis elegans примерно в 35 раз меньше ДНК чем у человека. Он имеет длину приблизительно 100 миллионов пар оснований и содержит приблизительно 20 000 генов. Большинство этих генов кодирует белки, но, вероятно, среди них есть примерно 1 000 генов РНК. Благодаря простоте генетического аппарата нематода C. elegans была первым многоклеточным организмом чей геном был полностью прочитан (Stein et al, 2003 ). Вышеизложенное сделало C. elegans одним из наиболее удобных объектов для исследования механизмов генетического контроля процессов морфогенеза при развитии и регенерации (Kimble 1981 ). В 2006 году двое исследователей Эндрю Файер (Andrew Fire) и Креиг Мелло (Craig C. Mello) были удостоены Нобелевской Премии по физиологии и медицине за разработку метода исследования генов с помощью, так называемых интерферирующих двухцепочечных РНК (double-stranded RNA interference) – dsRNAi (Fire et al., 1998 ). В 1998 году они показали, что введение двуцепочечной РНК в организм круглого червя Caenorhabditis elegans приводит к снижению активности гена мишени с ДНК комплементарной к данной дцРНКи. Результатом этой работы стало появление метода и термина РНК-интерференция (Boutros et al.,2004 ) . Синтетическую двуцепочечную РНК, комплементарную заданному гену, вводят в клетку или организм, где чужеродная молекула РНК запускает систему РНК-интерференции. Этот метод позволяет исследователям значительно снижать уровень экспрессии соответствующего гена. Изучение последствий снижения экспрессии интересующего гена позволяет выяснить физиологическую роль продукта данного гена-мишени. Так как система РНК-интерференции не может полностью выключить экспрессию гена, данный метод называется «нокдауном гена» — в отличие от полного удаления гена, «нокаута гена»( Voorhoeve , Agami 2003 ).
В дальнейшем был выяснен механизм этого явления. Оказалось, что экзогенная (чужеродная) двуцепочечная РНК запускает систему РНК-интерференции, активируя рибонуклеазу Dicer которая разрезает длинные молекулы дцРНК (dsRNA) на короткие фрагменты порядка 21-23 нуклеотидов, называемые siRNA. Одну из двух цепочек каждого фрагмента называют «направляющей», эта одноцепочечная РНК далее включается в состав РНК-белкового комплекса RISC (RNA-induced silencing complex). В результате активности RISC одноцепочечный фрагмент РНК соединяется с комплементарной последовательностью 3’концевого нетранслируемого участка молекулы мРНК и вызывает либо разрезание мРНК белком Argonaute, либо иным образом вызванное ингибирование ее трансляции. Экзогенная двуцепочечная РНК узнаётся и связывается специальными эффекторными белками (например, RDE-4 у Caenorhabditis elegans), усиливающими активность белка Dicer. Такие РНК-связывающие белки облегчают перенос siRNA к комплексу RISC. Путь инициации РНК-интерференции в клетке может быть усилен в результате синтеза «вторичных» siRNA на матрице «первичных» малых интерферирующих РНК с помощью РНК-зависимой РНК-полимеразы (англ. RNA-dependent RNA Polymerase, RdRP). Организмы отличаются по способности воспринимать чужеродные двуцепочечные РНК и использовать их в процессе РНК-интерференции. Эффекты РНК-интерференции у растений и Caenorhabditis elegans (но не у млекопитающих) могут наследоваться. Система РНК-интерференции является важной частью иммунного ответа к вирусам и к другому чужеродному генетическому материалу ( ). Показано что при вирусной инфекции у Caenorhabditis elegans повышается экспрессия белков Argonaute при этом черви, в организме которых повышается экспрессия генов белка Argonaute, активируют пути РНК-интерференции и приобретают устойчивость к вирусной инфекции (Lu et al.,2005 ,Wilkins et al., 2005 ). Предполагается что подобный механизм иммунитета может быть и у млекопитающих о чем свидетельствует тот факт, что некоторые вирусы содержат гены, снижающие ответ системы РНК-интерференции в клетках млекопитающих. Так, например, РНКi, экспрессируемые вирусом герпеса, могут вызывать образование гетерохроматина (конденсированного и поэтому неактивного хроматина) и приводить к переходу вируса в латентное состояние (Li et al .,2005 ). Caenorhabditis elegans является одним из наиболее часто используемых организмов-моделей для исследований методами функциональной геномики Это связано с тем, что:
- нематода C. elegans имеет простую анатомию, достаточно простой генетический аппарат и при этом геном ее полностью прочитан;
- эффекты сайленсинга (ингибирования активности от анг. Silence- заглушать) генов у нематоды наследуются;
- система RNAi у этого организма работает наиболее эффективно (Kamath, 2003 ). Доставка дцРНК в нематоду чрезвычайно проста. Нематодам можно скармливать клетки бактерий, например, Escherichia coli, содержащие требуемые дцРНК, которые при этом усваиваются через кишечник. Данный способ доставки РНК с пищей является эффективным с точки зрения эффективности сайленсинга генов и одновременно намного более дешёвым, простым и быстрым, чем инкубация червей в растворе, содержащем дцРНК или введение дцРНК в гонады (Fortunato, Fraser, 2005 );
- именно на C. Elegans разработана мощная и эффективная методика для безболезненного и высокоселективного ингибирования функций микроРНК in vivo (Zheng et al.,2010 , Kundu, Slack 2010 ).
У большинства других организмов доставка дцРНК связана с многочисленными сложностями и намного более трудоёмка. Тем не менее принимались попытки крупномасштабного исследования геномов и в культурах клеток млекопитающих (Cullen , Arndt 2005 )
Разработке метода РНК-интерференции предшествовало открытие в 1993 году у нематоды Caenorhabditis elegans эндогенно экспрессируемых (т.е. синтезируемых самим организмом) микроРНК (miRNA), которые способны избирательно подавлять экспрессию генов ( Lee, et al, 1993 ). Помимо C.elegans аналогичные миРНК были найдены практически у всех многоклеточных организмов (Carrington , Ambros 2003 ). Позднее выяснилось, что эндогенно экспрессируемые миРНК могут играть важную роль в пост-трансляционной регуляции синтеза белков (Selbach et al, 2008 , Baek et al, ) и процессах регуляции морфогенеза ( , Grillari, Grillari-Voglauer 2010 ) . Возможно миРНК участвуют также в регуляции процессов старения, что было показано в экспериментах с долгоживущими и недолго живущими мутантами C.elegans (Honjoh, Nishida 2011 ,Shin et al, 2011 ). Были найдены мутанты у которых была снижена активность одного определенного гена, что приводило к существенному увеличению продолжительности жизни организма. Это говорит о том, что существуют гены регулирующие продолжительность жизни. К числу таких генов, очевидно, относятся гены снижающие активность инсулин/IGF- подобной сигнализации и сигнализации TOR. Учитывая тот факт, что существует значительное сходство между процессами старения у C.elegans и у человека эти данные могу способствовать выявлению возможной роли miРНК в процессах биомаркировки возрастных изменений организмов и разработке способов борьбы с функциональными расстройствами, вызванными старческим дряхлением (Kashyap, 2011 ) Большинство миРНК представлено уникальным классом коротких РНК, синтезированных на не кодирующих белки последовательностях, так называемой «молчащей» ДНК. Гены миРНК встречаются как в виде единичных генов, так и в виде кластеров (групп) генов. Нередко они имеют свои промоторы (участки необходимые для узнавания РНК-полимеразой точки начала транскрипции) и другие регуляторные участки. Некоторые миРНК находятся на участке интронов (вставок которые не кодируют аминокислотную последовательность белка и после транскрипции вырезаются в ходе процессинга первичного транскрипта в мРНК) принадлежащих генам кодирующим белки. Вначале миРНК транскрибируется в виде длинных первичных транскриптов.Затем они подвергаются процессингу сначала в ядре, а затем в цитоплазме. Образовавшиеся в результате молекулы РНК длиной в 21-23 нуклеотида включаются в комплекс RISC, а затем взаимодействуют с комплементарными 3’концевыми нетранслируемыми участками молекулы информационной РНК. Это приводит либо к деградации РНК мишени, либо к ингибированию ее трансляции в белок. Важно отметить что одна и та же последовательность миРНК может воздействовать на многие мРНК мишени. Такая способность контролировать экспрессию множества генов делает миРНК ключевым звеном в регуляции различных жизненных процессов включая развитие, регенерацию, метаболизм, старение, и т.д. Так, например обширный класс эндогенных миРНК (endo-siRNA) называемых 22G-РНК необходим для поддержания нормальной работы генетического аппарата (Gu et al., 2009 ) и правильной сегрегации хромосом (Claycomb et al., 2009 ) Открытие регенерации аксонов у Caenorhabditis elegans позволило начать эксперименты по изучению механизма и причин, по которым такая регенерация возможна у Caenorhabditis elegans и невозможна у млекопитающих (Ghosh-Roy, Chisholm, 2010 ) . Оказалось, что регенерация аксонов подавляется внеклеточной пероксидазой PXN-2, входящей в состав внеклеточного матрикса. При ингибировании функции PXN-2 активируется регенерации аксонов (Gotenstein et al., 2010 )
Механизмы копирования утраченных частей организма при регенерации на примере плоских червей – планарий. Позиционная память. Роль стволовых клеток и апоптоза (программируемой гибели клеток) в регенерации
правитьКлючевые слова: планария; полярность организма; закон позиционной памяти; стволовые соматические клетки; унипотентные, мультипотентные, плюрипотентные. стволовые соматические клетки; необласты; хроматоидные тельца; микро РНК; бластема; анатомические пропорции; апоптоз; ростовые факторы; Wnt; бета-Катенин; Hh; GSK3 - гликоген синтез киназа 3; интерактивные РНК; механизм клеточной навигации; лимит Хайфлика (Hayflick limit); тератома; тератокарцинома; химерный эмбрион; программы морфогенеза; возрастное несоответствие.
Плоские черви, известные биологам под названием планария, обладают удивительной регенерационной способностью. Планарии имеют длину примерно 1 – 2 сантиметра. На спинной поверхности в области головы у них имеется пара глаз, тогда как на брюшной поверхности в центральной части тела у них выпирает трубчатая глотка (фаринкс) выполняющая роль рта и ануса. Нервная система их состоит из головного мозга (ганглия) и пары нервных цепочек. Внутреннее брюхо, состоящее из трех отделов, снабжает организм питательными веществами без посредства кровеносной системы, которая у планарий отсутствует (Рис.7). Планарии имеют диплоидный геном длиной примерно 800 Mb (800 миллионов пар оснований) расположенный на четырех хромосомах (Cantarel et al., 2008 ) Несмотря на достаточно сложную морфологию любой даже небольшой фрагмент планарии (кроме мозга и глотки), вырезанный на участке от глаз до хвоста может регенерировать за период, не превышающий двух – трех недель в полностью восстановленное животное (Рис.8). Новообразованное животное в начале, как правило, значительно меньше по размерам, но при этом повторяет пропорции исходного организма (Brondsted , 1969 ).
Важной характеристикой организмов является их полярность. Морфогенез организмов удобно описывать, пользуясь системой трех осей: AP переднее – задней: от головы к хвосту (anterior - posterior); DV дорзо-вентральной: от спины к животу (dorso-ventral) и ML медиолатеральной: от середины тела к бокам (medio-lateral) которую в случае конечностей заменяет ось PD проксимодистальная: от основания к кончику.
При регенерации планарий точно копируется как AP (передне-задняя) полярность организма (т.е. со стороны фрагмента изначально более близкого к голове образуется голова, а со стороны более близкой к хвосту образуется хвостовая часть), так и DV (дорзо-вентральная, «спина-брюхо») полярность. В то же время при продольном рассечении тела планарии происходит латеральная (боковая) регенерация. При всех направлениях регенерации на старых тканях идет восстановление недостающей половины тела).
Рис. 7. Планарии имеют достаточно сложную морфологию: на спинной поверхности в области головы у них имеется пара глаз, а на брюшной поверхности в центральной части тела у них выпирает трубчатая глотка (фаринкс).
Рис.8. Каждый фрагмент регенерирует в новое животное.
Головная часть фрагмента, как правило, регенерирует первой и только потом начинается регенерация хвостовой части. Время необходимое для регенерации головы при рассечении планарии на различных уровнях зависит от удаленности места разреза от головы (Рис.9). Быстрее всего регенерирует голова, если разрез делается у переднего конца червя и медленнее у заднего – хвостового (Dubois, 1949 ).
Рис.9 Время необходимое для регенерации головы при рассечении планарии на различных уровнях зависит от удаленности места разреза от головы.
Рис.10 А.- Существо именуемое «двуликим Янусом». Б.- «Двуликий Янус» не возникает, если нижний разрез сделать не сразу, а через некоторое время после первого.
Это очевидно свидетельствует о наличии полярно ориентированного градиента «голова-хвост». Обмануть эту полярность удается в том случае, если сделать очень тонкие поперечные ломтики в области «шеи» планарии. В этом случае в результате регенерации появляется очень странное существо именуемое «двуликим Янусом» в честь римского бога с двумя лицами (Morgan 1898 ). У этого существа с обеих сторон ломтика образуются головы, одна из которых там где ей и следовало быть, а другая в том месте, где должен был образоваться хвостовой отдел (рис.10).
«Двуликий Янус» не возникает, если нижний разрез сделать не сразу, а через некоторое время после первого, когда уже достигнут определенный уровень регенерации и голова, которая начинает развиваться на переднем конце ломтика становится способной оказать регулирующее влияние на формирование хвоста на противоположном срезе ломтика. (Goss, 1969 ),
Изучение последовательных этапов регенерации планарии показало, что вначале в месте разреза начинает вырастать образование, напоминающее по форме купол. Это образование, названное регенерационной почкой или бластемой (от греч. Blastos- росток) первоначально представляющее собой скопление внешне совершенно одинаковых на вид клеток, начинает расти и после нескольких последовательных стадий дифференцировки формируют те части тела, которые требуют восстановления (Reddien, Sanches Alvorado, 2004 ). Образованию бластемы предшествует повышение скорости деления необластов в ответ на ампутацию и их направленная миграция к месту ранения (Salo and Baguna, 1984 ).
Наряду с образованием бластемы идет и другой процесс наиболее хорошо заметный при регенерации планарий из особенно маленьких фрагментов – это перестройка оставшегося фрагмента, необходимая для воспроизведения прежних пропорций в меньшем масштабе. Например, головной фрагмент, полученный отсечением на уровне «шеи» не только формирует бластему, но и становится уже и удлиняется. При этом на нем образуется новый фаринкс (глотка) в месте, где его раньше не было, но где он должен быть в уменьшенной копии. Одновременно размеры глаз и мозга (церебрального ганглия) пропорционально уменьшаются в соответствии с новыми масштабами (Oviedo et al, 2003 ; Pellettieri et al, 2010 ).
Аналогичную перестройку можно наблюдать и при длительном голодании планарий, когда размеры животного могут уменьшиться почти в 20 раз. Показано, что уменьшение размеров животного как при голодании, так и при регенерации, связано с сокращением числа клеток, а не с уменьшением размеров клеток (Romero & Baguna, 1991 ; Oviedo et al, 2003 ). Примечательно, что анатомические пропорции сохраняются, несмотря на столь значительное изменение размеров животного и числа его клеток. При восстановлении питания размеры животного возвращаются в норму за счет увеличения числа клеток путем интеркаляции (вставки) недостающих клеток опять же с соблюдением анатомических пропорций.
Сокращение числа клеток при голодании или же связанной с регенерацией перестройкой тела планарии обусловлено апоптозом (генетически запрограммированной гибелью клеток) и автофагией (Gonzales-Estevez et al., 2007 ). Это сокращение необходимо для реорганизации в соответствии с изменившимися порядковыми номерами клеток в общем плане позиционной информации организма, координирующем его анатомические пропорции (Agata et al., 2007 ).
Регенерация планарии подчиняется закону позиционной памяти (Wolpert 1969 ). Клетки запоминают свою позицию в исходном организме и стремятся при регенерации окружить себя клетками с соответствующими позиционными номерами. При этом определенные клетки фрагмента превращаются в организационные центры которые выделяют морфоген. Этот морфоген, действуя на соседние клетки, заставляет их дифференцироваться в соответствии с командой, задаваемой градиентом концентрации морфогена (Slack, 1987 Agata et al 200740, Meinhardt,2009 ) Это вроде как человек, стоящий в очереди с номерками на руке. Если очередь разбежалась, то при ее восстановлении он вперед себя поставит того кто по порядку был впереди, а позади того, кто по порядку был позади. Если этому человеку завязать глаза и повернуть его на 180 градусов то он начнет строить очередь в обратном порядке, нарушая общий порядок. Нечто похожее происходит, если небольшой ломтик из середины тела планарии подсадить в его первоначальную позицию, но при этом повернуть на 180 градусов его дорзо-вентральную ориентацию. Тогда на теле планарии в месте, где был подсажен трансплантат, нередко формируются эктопические (от гр. ektopos – смещенный) добавочные головы или хвосты (Kato et al 2001 ,Carlson, 2007 ).
Каким образом изменяется позиционная идентичность клеток после ампутации, каким образом она скоординирована в соответствии с анатомическими и физиологическими запросами организма, с процессом регенерации пока не известно. Как будто неведомый скульптор, ваяя живое существо, не только перераспределяет, но и удаляет ненужный материал, чтобы воплотить то, что намечено генетической программой. Так же как и во времена Чарльза Дарвина, мы остаемся в неведении, каким образом клетки четко координируют свои действия, хотя исследования в этом направлении ведутся, и определенные успехи уже есть (Forsthoefel and Newmark 2009 , Eivers et al 2008 ). Какая клетка, какую роль будет выполнять? В каком месте, в какой позиции она будет находиться? Какой способ она использует для ориентации и навигации? В каком виде представлен общий целостный план развития, анатомии и морфологии организма? Как он записан?
Все что мы знаем это то, что ядро живой клетки содержит всю необходимую генетическую информацию для осуществления развития и функционирования организма. Образно выражаясь, ядро выполняет в организме ту же роль, что и винчестер с жестким диском в ПК на котором записана программа. В ходе дифференцировки в разных органах начинает работать та часть генетической программы, которая командует синтезом белков в этом конкретном органе или отдельной клетке. А вот как все это скоординировано в целостный организм, увы, не ясно. Как это ни поразительно, но клетки, строящие организм, не только точно вовремя перестают расти, делиться, и принимают разную форму, они дифференцируются, а порой даже отмирают, чтобы получилась необходимая пространственная структура (Friedl P., Gilmour D. 2009 ). Попробуем разобраться в том что известно о этих процессах на сегодняшний день.
Клетки тканей общаются с соседними клетками с помощью химических внешних сигналов называемых ростовыми факторами. Одни клетки выделяют эти факторы, а другие реагируют на них.
Одним из таких факторов является Wnt (Yazawa et al., 2009 ). Сигнальная система Wnt обычно активируется при регенерации, что очевидно связано с необходимостью повысить интенсивность пролиферации (размножения) стволовых клеток (Reya, Clevers . 2005 ). Когда клетки получают Wnt, они реагируют на него снижением скорости деградации клеточных белков, так как Wnt «заставляет» разрушающий белки фермент, известный под названием GSK3 - Glycogen Synthase Kinase 3 (гликоген синтез киназа 3), оказаться в изоляции, прочно связанным с внутренней стороной мембраны так называемых мультивезикулярных телец цитоплазмы (Taelman et al, 2010 ) . Известно, что фермент GSK3 ответствен за «запуск» деградации порядка 20% клеточных белков, и в частности бета-Катенина (β-Catenin) – белка, который играет важную роль в механизме ориентации позиционной памяти (Croce, McClay,2006 Gurley et al, 2008 )
Связывание WNT и некоторых других секретируемых элементов этой сигнальной системы с рецепторами стабилизирует "β-катенин деградационный комплекс", состоящий помимо GSK3 из таких компонентов как: Axin, CKI и APC (Gurley et al, 2008). С другой стороны активация корецепторов LRP, приводит к его разрушению. Когда β-катенин- ключевой внутриклеточный элемент этого сигнального пути, накапливается в цитоплазме и входит в ядро, он активирует транскрипционные факторы TCF, регулирующие экспрессию множества генов (Adell et al,2010 ).
Если увязать эти данные с данными о том, что распад MDR белков может ограничивать продолжительность жизни и способность к пролиферации стволовых клеток, не трудно представить себе механизм возникновения «бессмертных» (immortal) раковых клеток (которые, как известно (Reya, Clevers . 200570), интенсивно выделяют Wnt), а также объяснить парадокс видимого бессмертия планарий, которые в результате регенерации (при которой также интенсивно выделяется Wnt) омолаживаются (Child, 1913 , 1915 ).
Еще в 1825 году удалось провести эксперименты, демонстрирующие на планариях особенности механизмов регенерации.
Рис.11. На каждой из половинок «шеи» образуется своя голова и получается двухголовая планария Рис. 12. Если у планарии с двумя хвостами (1 и 3) или двумя головами (6 и 7) отрезать лишнее ( 2, 4, 6, 7) то на их месте лишние хвосты и головы не восстанавливаются. Вместе с тем отрезанные хвосты и головы регенерируют в нормальную планарию. Нормальная регенерация происходит и в том случае, когда после удаления лишнего отсекают и оставшуюся голову (8, 9) или хвост (5).
Если удалить голову животного и через переднюю часть тела – «шею» сделать продольный разрез до глоточного отверстия, расположенного в середине нижней брюшной поверхности, то через некоторое время в результате регенерации на каждой из половинок «шеи» образуется своя голова и получается двухголовая планария. (рис.11). Интересно отметить, что если отрезать у двухголовой планарии одну из двух голов, то она вновь не регенерирует. Точно также не регенерирует дополнительный хвост (Herbert et al,1926 ). Оставшаяся культя постепенно деградирует и образуется нормальная планария (Рис. 12.). Отрезанная голова или хвост также регенерируют в обычную планарию. Если отрезать одновременно обе головы выше «шеи», то обе и регенерируют. Неспособна к регенерации отделенная от тела глотка (фаринкс) и головной мозг (ганглий) отрезанный выше глаз (Gentile et al., 2011 ). В проведенных в 2008 году экспериментах было показано, что в том случае если подавлено выделение β-катенина-1 у планарии S. mediterranea (Smed-β-catenin-1), бластема при регенерации дифференцируется в голову, независимо от полярности регенерирующего фрагмента. Более того если погрузить фрагменты планарии в раствор, содержащий двухцепочечные интерактивные РНК - RNAi (не путать с информационной РНК), избирательно подавляющие активность генов-мишеней (Newmark et al.,2003 ) (в данном случае ген Smed-β-catenin-1), то при регенерации образуются две головы с головным мозгом и глазами расположенными как у Двуликого Януса.(Iglesias et al., 2008 ). Более того у интактного (неповрежденного) животного, которому незачем регенерировать, хвост начинает превращаться в голову (Gurley et al., 2008 , Petersen , Reddien 2008 ). Прямо противоположный результат дает избыточная активация «сигнала» Wnt /β- катенин (Wnt/β-catenin). При гиперактивации этого сигнала происходит образование хвоста из бластемы, которая предназначалась для образования головы (Gurley et al., 2008). Важно, что при этом, однако не происходит дальнейшего изменения AP полярности (Kobayashi et al .2007 ). Аналогичным образом действует и связанный с эволюционно консервативным рецептором ростового фактора фибробластов FGFR (fibroblast growth factor receptor) ген nou-darake, который ограничивает дифференцировку тканей мозга только в пределах переднего участка и не меняет общую AP полярность (Cebrià et al.,2002 ). Из этого следует, что механизмы, которые определяют осевую полярность, могут быть разобщены с теми, которые контролируют морфогенез и дифференцировку (Felix, Aboobaker 2010 ). В определении полярности регенерации участвует также сигнальный путь Хеджехог - Hedgehog (Hh) – который регулирует сигнализацию Wnt/β-катенина (Rink et al., 2009 , Yazawa et al., 2009 ). Понижение уровня сигнала Hh с помощью Hh интерактивной РНК (Hh RNAi) ингибирует регенерацию хвоста. Активация Hh пути с помощью Патчед РНКи (patched RNAi ) приводит к избыточному образованию wnt патчед-1 (wntP-1) и регенерации хвостов вместо голов с передней стороны (Petersen , Reddien , 2009 ). Одновременное ингибирование WntP-1 и гена Патчед (Патчед обычно ингибирует сигнал Hh) возвращает высокий уровень Hh и приводит к регенерации голов с обеих сторон. За дорзовентральную (спина – брюхо) полярность (DV) у планарий очевидно отвечает костный морфогенетический белок (bone morphogenetic protein - BMP). Инактивация сигнализации регулируемой этим фактором приводит к смене дорзовентральной ориентации, сопровождаемой удвоением нервных цепочек и головного мозга (Molina et al.,2007 , Orii , Watanabe et al.,.2007 , Reddien et al., 2007 ). В некоторых случаях при этом на спинной стороне даже образуется вторая планария сросшаяся с первой как Сиамские близнецы (Molina et al.,200755). Процесс дорзовентральной ориентации при регенерации, как у планарий, так и у позвоночных (например, у лягушек Xenopus), регулируется, как минимум, двумя сигнальными регуляторами: BMP и анти-дорзализационным морфогенетическим белком (anti-dorsalizing morphogenetic protein – ADMP). Это свидетельствует об эволюционной консервативности механизмов активации и ингибирования дорзовентральной ориентации при морфогенезе. Помимо BMP и ADMP в регуляции участвуют ингибиторы: хордин (chordin) и ноггин (noggin). У планарий семейство генов ноггин насчитывает порядка десяти членов, которые подразделяются на собственно гены noggin (nog) и ноггино-подобные noggin-like (nlg) гены, которые отличаются тем, что содержат небольшую аминокислотную вставку в функциональном домене. При этом, в то время как гены ноггин действуют как обычно путем ингибирования BMP, силенсинг гена nlg8 планарий активирует эктопический нейрогенез и повышает уровень вентрализирующей bmp(RNAi), т.е. продукт гена noggin-like содействует BMP, а не ингибирует действие BMP как продукт гена noggin (Molina, 2011 ) Помимо вышеописанных Wnt/β-катенина и Hh найдены и другие факторы ответственные за регуляцию полярности регенерации, что свидетельствует о необыкновенной сложности механизмов клеточной навигации при регенерации (Gurley et al., 2010 , Oviedo et al., 2010 ) We show that after amputation, a wounding response precedes rapid re-organization of the A/P axis. Furthermore, cells throughout the body plan can mount this response and reassess their new A/P location in the complete absence of stem cells. While initial stages of the amputation response are stem cell independent, tissue remodeling and the integration of a new A/P address with anatomy are stem cell dependent. We also show that WNT5 functions in a reciprocal manner with SLIT to pattern the planarian mediolateral axis, while WNT11-2 patterns the posterior midline. Moreover, we perform an extensive phylogenetic analysis on the Smed-wnt genes using a method that combines and integrates both sequence and structural alignments, enabling us to place all nine genes into Wnt subfamilies for the first time ( )
Удивительная способность планарии к регенерации связана с особыми клетками, называемыми необластами. Эти клетки являются стволовыми соматическими клетками (Adult Somatic Stem Cells – ASSC). ASSC это клетки, которые сами не являясь терминально дифференцированными (т.е. прошедшими специализацию до конца) предназначены для производства специализированных клеток. Они способны многократно делиться. При делении каждая дочерняя клетка стоит перед выбором: остаться стволовой клеткой, какой была родительская или встать на путь, необратимо ведущий к полной дифференцировке. Это отличает их от дифференцированных клеток, которые обычно сами делиться не могут. Те стволовые клетки, которые порождают только один вид дифференцированных клеток, называют унипотентными, те которые порождают несколько видов дифференцированных клеток мультипотентными, а способные дать начало целому организму – плюрипотентными. Необласты планарий во время регенерации способны дать начало не только всем типам соматических клеток необходимых для восстановления морфологии планарии, но даже зародышевым клеткам необходимым для полового размножения планарий (Sato et al 2006 , Wang et al 2007 ). Необласты составляют примерно 25-30 % всех клеток организма планарии (Shibata et al, 2010 ), что примерно в несколько тысяч раз больше чем число ASSC у млекопитающих. (Для сравнения: у человека, например, количество взрослых мезенхимальных стволовых клеток обеспечивающих постоянную замену твердых тканей организма к 15 годам не превышает одной – двух клеток на 100 тысяч, а к 60 годам падает до одной клетки на миллион (Caplan, 2008 )). Если облучить планарии рентгеновскими лучами то при определенной дозе облучения они теряют способность к регенерации, что связано с повреждением необластов (Wolf and Dubois, 1948 ). Способность к регенерации у облученных планарий удается восстановить инъекциями выделенных необластов (Kobayashi et al 2008 ). Это с одной стороны свидетельствует об отмеченной выше плюрипотентности необластов, а с другой говорит о том, что необласты способны реагировать на запросы организма. Особенностью необластов является увеличенное соотношение ядро- цитоплазма. В тонкой прослойке цитоплазмы можно различить помимо свободных рибосом и митохондрий еще и так называемые цитоплазматические хроматоидные тельца, представляющие собой округлые, плотные структуры, не окруженные какой либо мембраной. Хроматоидные тельца являются отличительной особенностью необластов. Их число и размеры сильно уменьшаются при дифференцировке. Можно поэтому предположить, что хроматоидные тельца поддерживают способность необласта оставаться ASSC (Solana et al.2010 ). Эти структуры являются РНП гранулами так как они разрушаются при обработке рибонуклеазой. Предполагается, что хроматоидные тельца необластов участвуют в посттранскрипционной регуляции. В этих механизмах принимают участие несколько РНК-связывающих белков и микро РНК (miRNA) хроматоидных телец (Friedländer et al, 2009 ). Роль микроРНК в этих процессах стала проясняться только недавно (Grillari, Grillari-Voglauer 2010 ). Показано что малые РНК участвуют в реализации регенеративной способности планарии Schmittea mediterrenea. Так «нокдаун гена» Smed-Ago1 с помощью РНК-интерференции приводит к резко выраженным нарушениям процесса регенерации и существенному сокращению популяции необластов, что, очевидно, свидетельствует об участии микроРНК в процессах регенерации и функционировании необластов.
То, что планарии легко регенерируют из маленьких кусочков, содержат большое количество ASSC (Handberg-Thorsager et al., 2008 ), очень легко размножаются в условиях лаборатории, содержат многие белки аналогичные человеческим белкам (Mineta et al., 2003 ), а геном их к настоящему времени достаточно изучен, делает их очень удобной моделью для исследований в области регенеративной биологии и медицины (Gentile et al., 2011 ). Кроме того планарии могут служить отличной моделью для исследования морфогенеза. Их удивительная способность восстанавливать при регенерации анатомические пропорции своего тела очень важна для развития биоинженерии тканей и органов (Saltzman 2007 , Adell et al,2010). То, что стволовые клетки планарий способны неограниченно делиться - в отличие от стволовых клеток человека, число делений которых ограничено так называемым лимитом Хайфлика (Hayflick limit (Hayflick, 1965 )) и не превышает 40-50 делений – делает их также удобной моделью для исследования причин возрастного снижения числа стволовых клеток у человека (Rodier, Campisi, 2011 ). Другое важное отличие стволовых клеток планарии от ASSC человека заключающееся в том, что они способны неограниченно делиться и при этом не превращаются в раковые клетки, может помочь в разгадке тайны ограничений регенерационной способности у человека. Планарии с легкостью регенерируют свой мозг, тогда как у человека регенерация даже одной нервной клетки невозможна (Sun & He, 2010 , Cebrià 2007 ). Инъекции необластов (Kobayashi et al 2008) не ведет к образованию у планарии тератомы – опухоли в которой представлено множество разновидностей дифференцированной ткани вперемешку со стволовыми клетками, которые продолжают делиться, и образуют нечто сходное с каллусом (см. раздел 1.2.). А у взрослого человека и других млекопитающих трансплантация плюрипотентных или эмбриональных стволовых клеток обычно приводит к образованию тератомы (Gutierrez-Aranda, 2010 ), которая затем может превратиться в злокачественную опухоль – тератокарциному (Martin 1980 ). Если, однако, поместить клетки тератокарциномы в ранний зародыш млекопитающего (на стадии бластоцисты), то они включаются в состав клеточной массы бластоцисты и из такого химерного (то есть состоящего из клеток от разных организмов) эмбриона нередко развивается нормальное химерное животное. Почти во всех органах и тканях таких животных часть дифференцированных клеток происходит из клеток тератокарциномы, которые совместно с клетками нормального происхождения участвуют в построении здорового организма (Papaioannou et al.,1978 ).
Рис. 13. Если поместить клетки тератокарциномы в ранний зародыш млекопитающего (на стадии бластоцисты), то они включаются в состав клеточной массы бластоцисты и из такого химерного (то есть состоящего из клеток от разных организмов) эмбриона нередко развивается нормальное химерное животное. Почти во всех органах и тканях таких животных часть дифференцированных клеток происходит из клеток тератокарциномы, которые совместно с клетками нормального происхождения участвуют в построении здорового организма (Papaioannou et al.,1978). Аналогичная трансплантация клеток тератокарциномы или эмбриональных клеток во взрослый организм неизменно приводит к развитию тератокарциномы. У планарий такой зависимости от возраста нет.
Очевидно, что взаимодействие ткани взрослых млекопитающих с эмбриональными стволовыми клетками приводит к «сбою» программы морфогенеза из-за возрастного несоответствия (обусловленного биомаркировкой возрастных изменений с помощью специальных микроРНК). В роли таких биомаркеров ( ) могут выступать так называемые онкогенные миРНК, такие как, например miR-19, и некоторые другие члены кластера miR-17-92 (Grillari et al., 2010 ), а также микро РНК miR-302–367 (Barroso-delJesus et al., 2010 ).
Исследование планарий позволит понять, почему у планарий, где такая программа достаточно простая такие «сбои» не возникают.
Глава. РЕГЕНЕРАЦИЯ У НАСЕКОМЫХ И ПОЗВОНОЧНЫХ ОРГАНИЗМОВ
правитьКлючевые слова: шаблон морфогенеза, регенерация, дрозофила, Данио-рерио, zebrafish, амфибии, саламандры, мексиканский аксолотль, бесхвостые амфибии, лягушки и жабы, метаморфоз, неотения, цыпленок, мышки.
Задумывались ли вы когда-нибудь над тем, что хотя на Земле живет несколько миллиардов людей, каждый из которых отличается своей только ему присущей внешностью, голосом, отпечатками пальцев, каждый из нас, тем не менее, построен и развивается по типовому шаблону - "Человек - Homo sapiens". Каждый из нас уникальный шедевр сложнейшей архитектуры, созданный великим прорабом по имени Морфогенез. О том, как работает морфогенез, большинство из нас не имеет ни малейшего представления. Но есть пытливые умы, которые пытаются разгадать секреты его искусства. Помочь ученым проникнуть в тайны морфогенеза может обыкновенная плодовая мушка - Дрозофила.
Всего 9 дней длится магия превращения яйца во взрослую муху. За это время крошечная клеточка - оплодотворенная яйцеклетка - строит из себя летательный аппарат, состоящий из многих миллионов частей (клеток), каждая из которых выполняет свою, именно ей предназначенную функцию, четко согласуя ее с общим планом действий. Волшебство, которому мог бы позавидовать джин из сказок про Алладина, мы можем исследовать, размножая дрозофилу. Многого для этого не надо: широкие пробирки с питательной средой, закрытые ватным тампоном, термостат чтобы поддерживать необходимую температуру и бинокулярный микроскоп. Не удивительно поэтому, что дрозофила стала одной из самых распространенных моделей для изучения тайн живой природы ( ).
Как не восхищаться удивительному богатству красок, сложности и симметрии узоров на крыльях у бабочек (Рис. 2.а.). А ведь это наглядный урок возможностей морфогенеза и его генетической регуляции. Изучение этих восхитительных художественных произведений природы, возможно, позволит открыть способы тканевой инженерии морфогенеза.( )
Рис. 2.а. Как не восхищаться удивительному богатству красок, сложности и симметрии узоров на крыльях у бабочек. А ведь это наглядный урок возможностей морфогенеза и его генетической регуляции. Изучение этих восхитительных художественных произведений природы, возможно, позволит открыть способы тканевой инженерии морфогенеза.
Другой распространенной моделью для изучения морфогенеза является аквариумная рыбка Данио-рерио (zebrafish), принадлежащая к роду Данио семейства Карповых. Эта пресноводная тропическая рыбка, имеющая яркий, полосатый рисунок, не случайно выбрана исследователями. Дело в том, что она обладает способностью при регенерации плавников восстанавливать не только форму и структуру, но и цветовой рисунок удаленного плавника. Наблюдение за восстановлением пигментных клеток этой рыбки позволило сделать немало важных выводов о механизмах регенеративного морфогенеза. Помимо хвостового и парных плавников Данио-рерио способны регенерировать поврежденную сетчатку глаз, поперечно перерезанные зрительные нервы и спинной мозг, а также сердечную мышцу после резекции до 20% желудочка, причем практически без образования рубцов. Эти удивительные способности к восстановлению своей анатомии, а также необыкновенная скорость, с которой эта регенерация происходит (7-15 дней для комплексного восстановления костей, эпидермиса, мышц и кровеносных сосудов; до 45 дней для восстановления цветового рисунка плавника) сделали Данио-рерио излюбленной моделью для изучения регенеративного морфогенеза. Важно также отметить, что эта рыбка быстро размножается в лабораторных условиях, операция по ампутации ее плавников не представляет трудности, а последовательности ее генома полностью прочитаны и истолкованы. Данио-рерио имеет 25 хромосом и ее геном состоит примерно из 1,5 х 10 миллионов пар оснований. (см. http://zf-health.org/information/factsheet.html )
Самой удивительной способностью к регенерации среди позвоночных обладает класс земноводных, иначе называемый классом амфибий. Он делится на две основные группы: хвостатые амфибии (саламандры) и бесхвостые (лягушки и жабы). Саламандры – группа амфибий, которые не теряют хвоста в процессе метаморфоза (гр. Metamorphosis), т.е. перехода из одной формы развития в другую, выражающейся в резком изменении строения животного. Некоторые виды саламандр способны к неотении (фр. Neotenie), т.е. к способности достигать половой зрелости и размножаться в стадии личинки. Особенностью неотенических амфибий является очень быстрая регенерация по сравнению со взрослыми животными. Так, если саламандре для того чтобы восстановить конечность требуется несколько месяцев, то у мексиканского аксолотля на это уходит всего 3-4 недели. Амфибий легко размножать в условиях лаборатории, их содержание не требует больших затрат. На них легко проводить микрохирургические операции. Раны их заживают очень быстро, не требуя наложения швов и особых хлопот по послеоперационному уходу. Результаты регенерации обычно видны уже через одну две недели (Song, Stocum 2010 ). Удобной и общедоступной моделью является также цыпленок. Для его выращивания из яиц требуется только термостат - инкубатор Классической моделью для изучения регенерации (главным образом из-за того что они также как и человек относятся к млекопитающим) являются мышки.
Регенерация тканей у винной мушки Drosophila
правитьКлючевые слова: компартмент, поликлон, граница компартмента, морфоген, гены, hedgehog, decapentaplegic, wingless, Engrailed, избирательная адгезия клеток, миозин II, Y-27632, Rho-киназы, трансдетерминация, гомеодомен, клетки mixer, JNK, c-Jun amino-terminal kinase, способ регуляции морфогенеза и перепрограммирования клеток, регенерация имагинальных дисков,
Среди насекомых способности к регенерации существенно различаются. Если таракан с легкостью регенерирует свои конечности (French V 1980 ), то взрослая винная мушка такой способностью не обладает (Marsh, Theisen ,1999 ). Тем не менее, винная, или как ее еще называют, плодовая мушка Drosophila melanogaster, служит очень удобным объектом для исследований морфогенеза при регенерации, так как ее генетика очень хорошо изучена, а способностью к регенерации обладают ее имагинальные диски. Геном дрозофилы состоит из порядка 132 миллионов оснований и приблизительно 13 767 генов ( )
Дрозофила проходит в своем развитии личиночную стадию (Рис 2.1). Организм взрослой мухи, называемый имаго, в основном образуется из определенных групп клеток, называемых имагинальными клетками, которые расположены в теле личинки обособленно и называются имагинальными дисками. На стадии куколки ткани личинки разрушаются, и из имагинальных дисков образуются ткани взрослого организма. Известно 19 дисков, один из которых расположен на средней линии тела, а остальные 18 лежат попарно по бокам личинки. Диски представляют собой эпителиальные мешочки в форме сплюснутых шариков. Внешне клетки разных имагинальных дисков различить невозможно, однако многочисленные эксперименты показали, что задолго до своей внешней дифференцировки эти клетки приобрели наследственную специализацию, определяющую путь их дальнейшего развития - они детерминированы (лат. determinatio — ограничение, определение). Клетки разных имагинальных дисков детерминированы по-разному, что предопределяет образование из них определенных структур, например, крыла или ноги или глаз (Рис.2.2).
Яйцо
Личинка
Куколка
Взрослая муха (имаго)
Рис.2.1 Схема развития дрозофилы от яйца до взрослой мухи
Рис.2.2. Имагинальные диски в личинке дрозофилы и образующиеся из них структуры взрослого организма (схематично).
На протяжении многих циклов деления, при отсутствии гормонального сигнала, необходимого для запуска дифференцировки, клетки остаются детерминированными, но не дифференцируются. Это позволяет размножать клетки, пересаженные из имагинального диска личинки, в брюшке взрослых мух, которые успели «проскочить» стадию, когда гормоны запускают их дифференцировку. Таким образом, брюшко взрослой мухи может использоваться как естественная культуральная камера.
В любое время из брюшка мухи можно извлечь культивируемые клетки имагинального диска и исследовать их детерминацию. Для этого такие клетки имплантируют личинке, и после ее метаморфоза опознают образовавшиеся из них структуры взрослой мухи, так как при метаморфозе они дифференцируются на новом месте в ту структуру, которая соответствовала их первоначальному положению (Рис.2.3),
Рис.2.3), Метод изучения детерминации клеток. Клетки можно тестировать,помещая их в личинку, сразу (вверху) или после размножения в брюшке взрослой мухи (внизу). В обоих случаях образуются структуры соответствующие детерминации клеток исходного диска
Взрослые имаго дрозофилы не способны регенерировать свои конечности, однако имагинальные диски способностью к регенерации обладают (Hadorn,1963 ).
В исследованиях проведенных в 60-е годы прошлого столетия было найдено, что
имагинальные диски могут регенерировать после их фрагментации или массовой гибели их клеток. Даже после уничтожения 50% клеток имагинального диска из него все же развиваются нормальные ткани, так как оставшаяся часть ткани способна восстановить популяцию клеток имагинального диска и таким образом компенсировать потери (Haynie, Bryant, 1977 ).
Эти регенераторные способности имагинального диска вкупе с многочисленными генетическими методами, разработанными специально для изучения дрозофилы, делают эту муху отличной моделью для изучения регенерации (Bergantiños et al 2010 , Repiso et al 2011 )
Как уже было отмечено, небольшие участки или даже отдельные клетки имагинального диска обычно остаются детерминированными для образования после метаморфоза характерных для этого участка структур взрослой мухи (Bryant, Schneiderman, 1969 , Bryant, 1975 ).
Клетки, полученные диссоциацией дисков, способны реагрегировать. При этом они специфично узнают соседние клетки со сходными «планами» детерминации - избирательно агрегируют именно с ними таким образом, чтобы воссоздать первоначальный шаблон ткани (Garcia-Bellido, 1966 )
Таким образом, имагинальные диски, как следует из вышеизложенного, являются жестко детерминированной системой клеток. Между тем наличие у имагинальных дисков способности к регенерации предполагает существование механизмов способных в какой-то мере снимать жесткие ограничения на детерминацию и возможность частично перепрограммировать эти клетки.
Действительно, было найдено, что в условиях культуры клетки имагинальных дисков иногда дифференцируются в структуры, отличные от тех, которые должны были образоваться из данного диска. Такая трансдетерминация представляет собой переход из одного наследуемого состояния в другое. Было показано, что трансдетерминацию претерпевают не отдельные клетки, а группы клеток (Hadorn, 1978 ). При этом трансдетерминированные клетки могут возвращаться в состояние исходной детерминации или какой-то иной детерминации. Число таких состояний детерминации ограничено и клетки выбирают одно из них. Частота некоторых типов трансдетерминации наблюдается чаще других. Например, гениталии чаще превращаются в антенну или ногу, нога чаще превращается в крыло (Рис.2.5). Явление трансдетерминации иногда наблюдается не только в регенерирующих имагинальных дисках, но и в процессе морфогенеза. Показано, что небольшая группа клеток, называемая mixer cells (смешивающиеся клетки) нарушая все вышеизложенные правила (1.- строго хранить детерминацию и 2.- не пересекать границ компартмента) способны внедряться в соседний компартмент. Такое поведение mixer клеток обусловлено перепрограммированием этих клеток, связанным с активацией синтеза белка гомеодомена кодируемого геном Engrailed, регулируемой сигнальным путем JNK (c-Jun amino-terminal kinase). (JNK – эволюционно консервативный фермент, играющий центральную роль в метаболизме и развитии нечувствительности к инсулину при диабете 2-го типа (Zhang et al 2011 )). Этот процесс перехода клеток mixer сопровождается стереотипной локальной интеркаляцией (внедрением) клеток, превращающей данный участок границы из напряженного в расслабленный в целях морфогенетических преобразований формы границы (Gettings et al 2010 ). Сигнальный путь JNK по видимому представляет фундаментальный способ регуляции морфогенеза и перепрограммирования клеток в процессах развития и регенерации так как он был найден и при регенерации имагинальных дисков (Lee et al 2005 ).
Рис.2.5. Относительная частота трансдетерминации различных имагинальных дисков. Длинными стрелками показаны частые направления трансдетерминации, короткими - более редкие, пунктиром очень редкие.
Интересно отметить, что, несмотря на то, что в брюшке дрозофилы можно размножать клетки имагинального диска детерминированного на образование ноги, никакой активации процессов регенерации ноги у этой дрозофилы не происходит. Очевидно, организм взрослой мухи не способен использовать эти клетки для регенерации. Здесь возможна аналогия с образованием тератомы у взрослых млекопитающих при введении им эмбриональных стволовых клеток. Как мы уже отмечали ранее (см. раздел 1.1.7.) клетки тератомы или тератокарциномы, помещенные в зародыш на стадии бластулы, успешно участвуют в образовании химерного организма, тогда как у взрослого организма такая пересадка вызывает образование злокачественной опухоли (Bradley et al.,1984 ). Учитывая то, что по данным цитогенетического анализа, тератокарциномы, как правило, не являются результатом мутации (van Berlo et al.,1990 ) - их образование, очевидно, связано именно со «сбоем» программы развития организма.
Эпигенетика на примере дрозофилы
правитьКлючевые слова: трансдетерминация, гомеотические гены, мутация antennapedia, динамическая память, эпигеном, нуклеосомы, позиционная информация, гомеобокс, гомеодомен, каскады генов, Hox-гены, кластер, коллинеарность генов, генетическая регуляция, белки группы, Polycomb (PcG), Trithorax (TrX), память клеточной идентичности, морфологическая регенерация, CTCF- связывающие участки.
В процессе развития из одной оплодотворенной клетки образуется великое множество самых разнообразных по морфологии и функциям клеток. Учитывая тот факт, что все эти клетки имеют один и тот же геном, т.е. содержат одну и ту же последовательность ДНК, надо полагать, что должна существовать какая-то информация в дополнение к геному для регуляции набора генов активных в конкретном типе клеток ( , ). Изучение трансдетерминации привело к предположению, что существуют гены, выполняющие роль коммутатора, включающего формирование стандартного признака в ответ на определенные сигналы (Фурман и др. 1975 ). Характерная особенность таких генов, названных гомеотическими, состоит в том, что нарушение их функционирования приводит к формированию сложной альтернативной структуры, а не к образованию аморфной тканевой массы. Например, мутация antennapedia приводит к тому, что целая группа клеток в норме образующих антенну, коренным образом изменяет свое поведение и формирует ногу, в результате чего появляется муха с растущей на голове ногой (Рис 2.4.)
А - Голова нормальной мухи
Б - мутация antennapedia - муха с растущими на голове ногами вместо антенн
Трансформированные клетки ноги образуют участки соответствующие положению этих клеток в антенне (это соответствие показано красными стрелками). В гомологичных органах, очевидно, используется одна и та же система позиционной информации, однако клетки интерпретируют ее по-разному в зависимости от своей предыстории.
Рис. 2.4
Такого рода данные привели к идее о существовании специальной многоуровневой системы триггеров (переключателей) контролирующей дифференцировку клеток. Триггер является простейшим вариантом динамической памяти в отличие от статической памяти, каковой является последовательность ДНК генома (Корочкин 1977 ). Определенное состояние триггера может передаваться по клеточной линии (как бы наследоваться), поддерживая, таким образом, клеточную дифференцировку. Такая многоуровневая динамическая память была обозначена термином эпигеном (Ратнер 1975 ). В настоящее время изучением эпигенома занимается новая наука эпигенетика (греч. Επί - над, сверх и слово генетика) которая изучает наследуемые изменения фенотипа (внешних проявлений ), а также генетической активности, вызванных механизмами, не меняющими последовательностей ДНК генома ( ). Существование триггеров в эпигеноме, предполагает асимметричное не случайное распределение состояния триггера при делении клеток, благодаря которому первоначальный тип клетки дает начало двум новым клеткам: одной наследующей новое состояние триггера и одной наследующей материнское первоначальное состояние (Рис.2.4.). В связи с этим следует отметить, что во время репликации ДНК в состав хроматина одной дочерней спирали ДНК входят нуклеосомы, сформированные материнскими гистонами, тогда как другая дочерняя спираль ДНК содержит только новые гистоны, синтезированные параллельно с синтезом ДНК. Об этом свидетельствует образование свободной ДНК лишь с одной стороны репликативной вилки при блокаде синтеза гистонов циклогексимидом ( ) Для развития многих типов дифференцированных клеток по схеме эпигенома требуется комбинация сравнительно небольшого числа ключевых регуляторных белков ( ). Следует отметить, что реакция клеток на один и тот же белок – активатор различается в зависимости от того какой путь развития уже прошла данная клетка. Так, например, благодаря вышеописанному динамическому механизму клеточной памяти, попав в поле деятельности градиента концентрации морфогена, клетка интерпретирует одну и ту же позиционную информацию по разному (в зависимости от своей предыстории) и соответственно своей интерпретации реагируют на нее. Например, в случае мутации antennapedia Дрозофилы сигналы указывающие положение клеток в придатке одинаково эффективны как по отношению к клеткам «решившим» стать клетками антенны, так и по отношению к ним, когда они «выбрали» путь, ведущий к образованию ноги. В результате трансформированные клетки образуют те участки ноги, которые соответствуют положению этих клеток в антенне ( ) (Рис. 2.4.). Гены Antennapedia являются примером генов содержащих гомеобокс - последовательность ДНК, обнаруженную в генах, вовлеченных в регуляцию развития у животных, грибов и растений. Гомеобокс состоит приблизительно из 180 пар нуклеотидов, он кодирует белковый домен длиной в 60 аминокислот (гомеодомен), который может связывать ДНК. Гены, содержащие гомеобокс, кодируют факторы транскрипции, которые, как правило, переключают каскады других генов. Гомеодомен связывается с ДНК специфическим образом. Однако специфичности одного гомеодомена, как правило, не хватает для распознавания целевого гена. Обычно белки, содержащие гомеодомен, действуют в промоторной области целевых генов в комплексе с другими факторами транскрипции, часто также белками, содержащими гомеодомен. Такие комплексы, как правило, имеют намного более высокую специфичность по сравнению с белком, имеющим единственный гомеодомен. Наиболее изученными и наиболее консервативными белками, содержащими гомеодомен, являются Hox-гены, которые контролируют сегментацию во время развития. Кроме того Hox-гены играют ключевую роль в эмбриональном развитии животных, управляя дифференцировкой отделов тела вдоль передне-задней оси. Эти гены отличаются уникальным поведением. Обычно они располагаются группами (кластерами), линейный порядок генов внутри кластера обычно соответствует времени работы гена в развитии. Данное явление называют коллинеарностью ( ).Показано, что существует две основные фазы в регуляции генов Hox : Первая - где непосредственно расписано время активации конкретных генов (эта фаза затрагивает глобальные регуляторные воздействия на гены расположенные вне кластера Hox), и вторая - где регуляция затрагивает взаимоотношения генов внутри кластера. Это свидетельствует о существовании разных типов коллинеарности, функции которых могут различаться ( ). Важную, но пока непонятную роль в работе генов кластера Hox по видимому играет белок CTCF связывающийся с последовательностью CCCTC на границе кластера с другими генами. Показано, что хотя отсутствие этого белка на ранних стадиях развития никак не отражается на работе генов, на более поздних стадиях делеция Ctcf вызывает массивный апоптоз приводящий к почти полной потере структур конечностей ( ). У дрозофилы известны две группы генов играющих ключевую роль в процессах развития. Это гены, кодирующие белки группы Polycomb (PcG) и Trithorax (TrX) Белки TrX функционируют в качестве активаторов транскрипции, тогда как гены PcG работают в качестве глушителей активности генов. Ответственные за регулировку этих белков факторы могут запоминать и на протяжении многих поколений клеток поддерживать активное или пассивное состояние конкретных генов, несмотря на то, что активатор или репрессор, вызвавший это состояние, давно исчез. Таким образом, система TrX / PcG может играть роль памяти клеточной идентичности (Ringrose, Paro 2007 ). В качестве факторов носителей клеточной памяти могут выступать регуляторные элементы генов Hox. Предполагается, что регуляция генов гомеобокса посредством системы TrX / PcG играет важную роль в образовании и поддержании механизмов позиционной информации необходимой для осуществления точной реконструкции первоначальной структуры путем морфологической регенерации (Tamura 2010 ). Bernd Schuettengruber, Anne-Marie Martinez, Nicola Iovino & Giacomo Cavalli (December 2011) Trithorax group proteins: switching genes on and keeping them active Nature Reviews Molecular Cell Biology 12, 799-814 | doi:10.1038/nrm3230
Abstract
Cellular memory is provided by two counteracting groups of chromatin proteins termed Trithorax group (TrxG) and Polycomb group (PcG) proteins. TrxG proteins activate transcription and are perhaps best known because of the involvement of the TrxG protein MLL in leukaemia. However, in terms of molecular analysis, they have lived in the shadow of their more famous counterparts, the PcG proteins. Recent advances have improved our understanding of TrxG protein function and demonstrated that the heterogeneous group of TrxG proteins is of critical importance in the epigenetic regulation of the cell cycle, senescence, DNA damage and stem cell biology.
Представления о границе тканевых компартментов на примере дрозофилы
правитьКлючевые слова: морфогенез, клон, компартмент, поликлон, граница компартмента, градиент, морфоген, белок, hedgehog, decapentaplegic, wingless, избирательная адгезия клетки, миозин II, натяжение между клетками, Y-27632, Rho-киназа, трансдетерминация, mixer cells, смешивающиеся клетки, внедрение, интеркаляция, белок гомеодомена, ген еngrailed, сигнальный путь, JNK, c-Jun amino-terminal kinase, нечувствительность к инсулину, перепрограммирование клеток, регенерация имагинальных дисков.
Изучение детерминации клеток у дрозофилы с помощью мутантных клонов (от англ сlon - тканевая культура генетически идентичных клеток ), полученных с помощью рентгеновского облучения, показало что крылья и другие органы состоят из ряда участков «компартментов». Отношения между клонами удобно изучать на широких и плотных крыльях дрозофилы. Пятна образованные маркированными клонами обычно располагаются на крыле случайным образом и имеют, как правило, довольно неправильные очертания. Это говорит о том, что потомки одной клетки могут распределяться в пространстве (плоскости крыла) по-разному – их расположение не детерминировано. Однако клоны вблизи центральной оси крыла как бы разделены невидимой границей между передней и задней частями крыла, проходящей у всех мух одинаковым образом. Клоны могут быть расположены с той или другой стороны от этой границы, но, как предполагалось до недавнего времени, никогда ее не пересекают. В тех местах, где клоны примыкают к этой границе, края соответствующих пятен бывают необычно четкими и прямыми (Crick, Lawrennce, 1975 ). Участок, отделенный такого рода границей называемый компартментом, является поликлоном т.е. состоит из различных клонов. Интересно отметить, что эта граница не совпадает с центральной жилкой крыла (Рис.2.4 ).
Рис.2.4. Даже если один из клонов компартмента растет быстрее остального крыла, он все равно не переходит границу компартмента. Эта граница не совпадает с центральной жилкой крыла или каким-либо видимым механическим барьером, ограничивающим рост клона.
Вместе с тем, даже если один из клонов компартмента растет быстрее других и поэтому занимает большую его часть, вытеснив другие клоны, он все равно, как правило, не переходит границу компартмента. Быстро растущий клон может заполнить почти весь свой компартмент и, тем не менее, размеры и строение такого компартмента не отличаются от нормы. Показано, что размеры компартмента у дрозофилы определяются не детерминированным числом делений клеток, а пространственным сигналом, извещающим клетки о том, что структура, в пределах которой они растут, достигла нормальных, предопределенных «планом», размеров (Simpson, Morata.1981 ). Каким образом достигается столь четкая и скоординированная в соответствии с планом архитектуры крыла граница пока остается загадкой. Предполагается, что такой сигнал возникает в результате межклеточных взаимодействий и зависит от расстояний, разделяющих клетки. Конечности Drosophila подразделяются на передний и задний компартменты. Предполагается, что клетки обоих компартментов растут, подчиняясь локальной концентрации морфогена. В качестве такого морфогена был предложен секретируемый клетками заднего компартмента белок hedgehog, который в свою очередь индуцирует в соседствующих с ними клетках переднего компартмента секрецию белка decapentaplegic или же белка wingless (Basler, Struhl 1994 ). Была высказана гипотеза, которая объясняет четкость и прямоту границы между компартментами избирательной адгезией: клетки одного поликлона предпочитают образовывать связи друг с другом, а не с соседним поликлоном (Wright, Lawrence, 1981 ) . Такое поведение объясняется двумя направленными друг к другу градиентами двух морфогенов, управляющих адгезией клеток. Важную роль в механизме такой избирательной адгезии, очевидно, выполняет немышечный сократительный белок миозин II. Показано, что вдоль границы компартментов натяжение между клетками повышено в 2,5 раза по сравнению с остальной тканью. Это натяжение ослабляется в присутствии Y-27632 – препарата, который является ингибитором Rho-киназы (Routier et al 2010 ) - главного активатора миозина II. Очевидно, локальное повышение натяжения связей между клетками, регулируемое посредством миозина II, является тем механизмом, который направляет и определяет выпрямленную «по струнке» и четкую границу между компартментами (Landsberg et al 2009 ).
Крылья бабочек – уроки мастерства морфогенеза
правитьКлючевые слова: симметрия узоров, генетика морфогенеза, полиморфизм, гинандроморфные бабочки, мозаичный гинадроморфизм, локус HmYb бабочек Heliconius.
Почти у всех бабочек рисунок на левой паре крыльев идентичен рисунку правой, являясь его зеркальным отражением. Расположение различно окрашенных чешуек на крыле закономерно и образует сложные рисунки, характерные для каждого вида, а часто и систематической группы.( )
Появление различных форм, отличающихся по окраске и ряду признаков, внутри одного вида называемое полиморфизмом, дает богатый материал ( ) для изучения особенностей генетики морфогенеза ( ) (Рис. 2.6. )
Каждая пигментная чешуйка содержит в себе только один пигмент ( ). Наиболее часто это меланин, придающий чёрный и коричневый цвета, и липохромы каротиноиды — характеризующиеся светочувствительностью и обуславливающие жёлто-зелёную, жёлтую, бурую, оранжевую и красную окраску. При оптической окраске в чешуйках бабочек возникает тонкослойная интерференция. Пигмент в нижней части этих чешуек не пропускает свет и придаёт бо́льшую яркость интерференционной окраске.
Лучи света, проходя через чешуйки, отражаются как от их внешних, так и от внутренних поверхностей, и в результате два отражения накладываются и усиливают друг друга ( ). Таким образом, цветные узоры специфичные для каждого вида образуются как результат целого комплекса генетически запрограммированных различных проявлений морфогенеза ( ). Это и изменения толщины мембран, и пигментация, и жилкование, и распределение волосков.
Рис. 2.6. Варианты полиморфизма Чешуекрылых
По материалам: ru.Wikipedia.org
Сезонный полиморфизм Географический полиморфизм Половой диморфизм
Пестрокрыльница изменчивая (Araschnia levana). Вверху весеннее поколение Araschnia levana f. levana. Внизу летнее поколение Araschnia levana f. prorsa
Самец Ornithoptera priamus urvillianus (верхнее фото).
Самец Ornithoptera priamus poseidon (нижнее фото).
Ornithoptera croesus. . Самец (наверху) и самка.
Во время развития бабочки возможны генетические изменения, приводящие к появлению гинандроморфных бабочек — полусамцов-полусамок, у которых одна сторона тела полностью мужская, а другая — женская (так называемые билатеральные гинандроморфы). Иногда аномальными становятся лишь небольшие участки тела бабочки, тогда говорят о мозаичном гинадроморфизме. Так, в различных участках крыла у самца могут появиться участки окраски самки и наоборот. Как правило, такие особи нежизнеспособны, но при сохранении функциональности внутренних органов имаго являются жизнеспособными. У чешуекрылых основой для появления гинандроморфа обычно является образование двуядерных ооцитов, у которых одно из ядер образуется, например, из ядра полярного тельца. Если одно из этих ядер содержит Z-хромосому, а другое — W-хромосому, то при оплодотворении двумя спермиями при делении этих ядер могут возникнуть клетки-потомки мужской и женской половин тела.
Рис 2.7. Гинадроморфизм Во время развития бабочки возможны генетические изменения, приводящие к появлению гинандроморфных бабочек — полусамцов-полусамок, у которых одна сторона тела полностью мужская, а другая — женская (так называемые билатеральные гинандроморфы)
Гинандроморф парусника Papilio glaucus. Левая половина тела - самца, правая - самки. Фото из ru.Wikipedia.com
Интересные данные для понимания механизмов морфогенеза получены при изучении процессов образования пятна – «глазка» на крыльях. Пигментированные концентрические круги глазка образуются в ходе развития из так называемого фокуса состоящего примерно из 300 клеток в центре намечающегося глазка. Еще в 1980 году было обнаружено, что если фокус взятый у особи в фазе кокона пересадить с одного места будущего крыла на другое он может индуцировать образование эктопического (находящегося не на своем месте) глазка в окружающих его клетках ( ). Более того, прижигание фокуса ингибирует образование глазка. Эти эксперименты позволили предположить, что клетки фокуса секретируют сигнальные молекулы, которые действуют как морфоген. Клетки окружающие фокус получают разные количества морфогена и в зависимости от концентрации морфогена в зоне где находится та или иная клетка, она активирует синтез того или иного пигмента ( ), что приводит к образованию узора из концентрических цветных колец (рис. ). В последующих исследованиях на роль таких морфогенов были предложены самые различные сигнальные молекулы ( , , , ) что, очевидно, свидетельствует о том, что реальный механизм морфогенеза глазков пока не совсем понятен ( ). Интересно отметить, что ранение крыла на стадии кокона индуцирует образование в месте ранения эктопических глазков ( ). Вызванные ранением глазки, активируют те же гены, что и нормальные вызванные фокусом ( ), что свидетельствует о том, что независимо от природы инициирующего сигнала, последующее развитие событий в сети генетических сигналов, регулирующих образование пигментных клеток глазка идет по «проторенной дорожке» ( ) (Рис.). Анализ микроРНК бабочек Heliconius показал, что в районе локуса HmYb кодирующего желтую окраску крыльев меняются главным образом две миРНК и обе они связаны с развитием, а не с морфологией цветовых узоров ( ). Таким образом, до настоящего времени не ясно как регулируется морфогенез цветовых узоров.
2.5. Регенерация органов у рыбки Данио-рерио (zebrafish)
Ключевые слова: Данио-рерио, zebrafish, регенерация, механизмы регенеративного морфогенеза, бластема, прогениторные клетки, пропигментные клетки, сетчатка глаз, зрительные нервы, спинной мозг, сердечная мышца, пролиферация, кардиомиоциты, ген fgf20a, белки JunB, фосфорилирование, JNK , Jun N-концевая киназа, Jun N-terminal kinase,
Аквариумная рыбка Данио-рерио (zebrafish), принадлежащая к роду Данио семейства Карповых является пресноводной тропической рыбкой. Ее длина не превышает 4-5 см. Она имеет яркий, полосатый рисунок, чем-то напоминающий зебру, за что она и получила свое английское название zebrafish. Уникальной способностью этой рыбки при регенерации плавников восстанавливать не только форму и структуру, но и цветовой рисунок удаленного плавника уже давно заинтересовались исследователи морфогенеза. Наблюдение за восстановлением пигментных клеток этой рыбки позволило сделать немало важных выводов о механизмах регенеративного морфогенеза. После ампутации плавника на раневой поверхности появляется лишенная всякой окраски бластема. Вслед за этим, от места отсечения, начинают отрастать костные лучи плавника. Однако даже через 3 недели после ампутации, когда общая структура плавника уже восстановлена, на нем все еще видна только одна из трех (красной, черной и желтой) цветных полос. Восстановление характерной окраски плавника происходит лишь позднее, через 40-50 дней после ампутации. Таким образом, пигментные клетки восстанавливаются путем дифференциации из бесцветных прогениторных (в данном случае пропигментных) клеток (Goodrich, Green, 1959 ). Важно отметить, что при этом восстанавливается не только трехцветная окраска плавника, но и фактически равная исходной ширина каждой полосы. При регенерации хвостового плавника Данио-рерио наблюдается целый комплекс процессов восстановления - восстанавливаются: эпидермис, кости, мышцы, кровеносные сосуды, нервы.
Помимо хвостового и парных плавников Данио-рерио способны регенерировать поврежденную сетчатку глаз, поперечно перерезанные зрительные нервы и спинной мозг, а также сердечную мышцу после резекции до 20% желудочка, причем практически без образования рубцов.
Удивительно, что при регенерации сердца, образовавшиеся новые сердечные клетки способны функционально интегрироваться в поврежденную, но продолжающую непрерывно сокращаться сердечную мышцу. В связи с этим необходимо отметить, что в отличие от имеющих четырехкамерное сердце и два круга кровообращения, млекопитающих (которые способностью регенерировать сердце обладают только до раннего постнатального периода) у рыб сердце двухкамерное и только один круг кровообращения. Кроме того, кардиомиоциты у взрослых млекопитающих не способны делиться. Даже при гипертрофии, когда количество ДНК в сердце увеличивается, это увеличение идет за счет полиплоидии, а не увеличения числа клеток. Что же касается мышечных клеток сердца рыбы, то они не теряют способности к делению и при повреждении начинают активную пролиферацию. Причем пролиферируют не только клетки поврежденного участка, но и клетки всего сердца.
Изучение мутаций у Данио-рерио показало, что в инициации процесса регенерации хвоста важную роль играет ген fgf20a, активация которого наблюдается вскоре после ампутации ( ). У млекопитающих подобный ген участия в регенерации не принимает. Помимо гена fgf20a для нормальной регенерации плавника необходимы белки JunB, которые во время регенерации хвостового плавника фосфорилируются JNK - Jun N-концевой киназой ( Jun N-terminal kinase ) ( ). Белок JunB является членом семейства транскрипционных факторов AP-1 (активаторный белок – 1) (activator protein-1), участвующих в регуляции клеточной пролиферации ( ). Предполагалось, что он ингибирует пролиферацию клеток и их трансформацию в раковые клетки, регулируя транскрипцию генов кодирующих белки промоторы клеточного цикла, такие как циклин D (CyclinD) или p16INK4A, а также циклин A (CyclinA) или p19(ARF). У человека, в отличие от рыб, этот белок не содержит участка для фосфорилирования ( ) и может играть роль онкогена вызывающего лимфому ( ).
Важную роль в механизме регенерации плавника может играть реактивация молчащих эмбриональных генов. Такая реактивация генов может быть связана с деметилированием гистона Н3. Об этом свидетельствует ингибирование процесса регенерации хвостового плавника в случае ингибирования деметиллазы
me3K27 H3 (me3K27 H3 demethylase), так как в результате не смог активироваться промотор одной из мишеней этого фермента (dlx4a – ген гомеобокса из семейства генов участвующих в формировании придатков в процессе эмбриогенеза).
Регенерация конечностей у земноводных
правитьНесмотря на то, что клеточные и молекулярные механизмы, участвующие в развитии конечностей у четвероногих животных высоко консервативны, регенеративные способности конечностей после ампутации у разных видов существенно различаются. Это очевидно связано с тем, что после ампутации создаются условия совершенно отличные от тех, в которых первоначально развивался удаленный орган. Так, например, представители амфибий – тритоны и саламандры способны полностью восстановить свои конечности (Wallace (1981 , Stocum,2004 ). Лягушки обладают разной степенью регенеративной способности в зависимости от стадии их жизненного цикла. Если ранние головастики Xenopus laevis и способны к полной регенерации конечностей и хвоста (за исключением переходного «рефрактерного» периода, во время которого хвост не регенерирует), то в последующем их регенеративная способность прогрессивно падает в ходе дальнейшего развития ( ). В результате этого если на ранней стадии после метаморфоза Xenopus laevis еще способен образовывать гипоморфные (недоразвитые) регенераты конечностей, то в последующем даже эта способность полностью исчезает ( ). У млекопитающих и у птиц регенеративные способности конечностей намного ниже. Тем не менее, у человека и у некоторых других млекопитающих возможна регенерация кончиков пальцев на стадии эмбриона и в первое время после рождения ( , ). Регенерация пальца у человека и мыши происходит лишь в том случае, если ампутации подвергся периферический (дистальный) участок концевой фаланги и если рана остается открытой, а не закрыта путем зашивания кожи над раной. После ампутации на более проксимальном уровне, новорожденная мышь не способна регенерировать утраченную часть, и происходит гипертрофия поврежденной кости ( , ). До недавнего времени предполагалось, что в регенерации конечностей задействованы главным образом стволовые клетки, однако оказалось, что это не так. Оказалось, что бластема формируется после закрытия раны путем дедифференциации (частичной потери специализации) по меньшей мере трех терминально (окончательно) дифференцированных типов клеток: фибробластов ( ), кератиноцитов ( ), и миофибриллярных клеток ( ). Кроме того в формировании бластемы, по- видимому, участвуют резервные миобласты (сателлитные стволовые клетки мышцы), но не так существенно как считалось ранее ( ). Несмотря на столь гетерогенное (разное) происхождение клеток бластемы, они под микроскопом выглядят как однородная популяция клеток. Лишь недавно их перестали считать единым типом клеток ( ). В связи, с чем сразу возникли вопросы не только о происхождении различных детерминированных (термин см. 1.2.1.) клеток бластемы, но и о механизмах, которые, после повреждения тканей, организуют: 1. высвобождение этих клеток из различных тканевых структур; 2. их миграцию к месту образования бластемы; 3. их дедифференциацию (потерю дифференцировки) с сохранением детерминации в так называемые прогениторные (progenitor) клетки; и 4. их скоординированное в соответствии с шаблоном распределение в бластеме и рост ( , ). Механизм дедифференцировки клеток привлекает особое внимание исследователей в связи с тем, что его изучение и использование для терапии сулит значительный прорыв в современной медицине. Интенсивные исследования способов перепрограммирования клеток позволили получить индуцированные плюрипотентные стволовые клетки – ИПСК (iPSC) , аналогичные эмбриональным стволовым клеткам –ЭСК (ЕSC). Было установлено, что линии ИПСК отличаются друг от друга, поскольку такие стволовые клетки хранят «воспоминания» о своём прошлом ( ). Прогениторные клетки бластемы отличаются как от эмбриональных стволовых клеток, так и от ИПСК тем, что хотя они и дедифференцированы, но не до такой степени плюрипотентности. Эмбриональные стволовые клетки, а также ИПСК можно использовать для клонирования целого организма, но они непригодны для бластемы, задача которой восстановить конкретную часть организма (руку или ее часть, ногу, сетчатку глаза, зуб и т.д.). Дедифференциация происходит в сочетании с высвобождением клеток из их окружения в результате лизиса (разрушения) связывающего их тканевого матрикса. Показано, что дедифференцированные клетки синтезируют характерные для их состояния факторы, такие как, например: Nrad ( ), msx1 ( ), rfrngand notch ( ). Вместе с тем синтезируются факторы необходимые для перепрограммирования клеток. Так, например, активируется синтез транскрипционного фактора Lin 28, ген которого в сочетании с генами Oct4, Sox2 и Nanog используется для перепрограммирования фибробластов в ИПСК ( ). Интересно отметить, что Lin28 кодирует цитоплазматический мРНК – связывающий белок ( ), который, связываясь с мРНК кодирующей инсулиноподобный ростовой фактор 2 (Insulin-like growth factor 2 - IGF2) активирует ее трансляцию ( ). Кроме того Lin28 блокирует созревание let-7 микроРНК в эмбриональных стволовых клетках мыши ( ). Показано, что при регенерации повышается синтез факторов плюрипотентности (Sox2, Klf4, и c-myc) в регенерирующих конечностях и хрусталике тритона ( ). У аквариумной рыбки Данио-рерио (zebrafish), по крайней мере, два таких фактора - (Oct4 и Sox2) необходимы для нормальной регенерации хвостового плавника ( ). Помимо активации определенных генов, для регенерации необходимо создать особые условия. Созданием этих условий занят эпидермис покрывающий рану. При исследовании интенсивности синтеза различных белков в регенерирующей конечности аксолотля обнаружено, что активнее всех белков в бластеме синтезируется синтетаза окиси азота (neuronal nitric oxide synthase- NOS1) – белок, идентифицированный как фермент синтезирующий окись азота NO – газ, обладающий широким спектром биологического действия. Активность синтеза NOS1 превышала синтез других белков с первого по 7-й день после ампутации конечности ( ). Ген, кодирующий NOS1 активируется также в культе конечности Xenopus на стадии когда регенерация возможна, но не активен на стадии когда способность к регенерации отсутствует ( ). Иммунное окрашивание показало, что NOS1 синтезируется главным образом в эпидермисе бластемы в период ее формации. Такая локализация NOS1 свидетельствует о том, что NO (являющийся индифферентным газом способным быстро диффундировать и свободно проникать через плотные клеточные слои и межклеточное пространство) диффундируя из эпидермиса вглубь бластемы и связываясь там с кислородом воздуха (замечательное свойство NO непосредственно связываться с кислородом воздуха, образуя двуокись азота) может поддерживать состояние гипоксии и, таким образом имитирует в бластеме условия приближенные к ранним стадиям развития зародыша. Такие условия, очевидно, необходимы для контроля индуцируемых гипоксией факторов – ГИФ (HIF) (Brune et al 2003 ), которые играют важную роль в регуляции способности клеток к дедифференцировке и пролиферации ( , ). Помимо этого окись азота, обладая бактерицидными свойствами, поддерживает стерильность в ране ( ). Однако главное что дает NO регенерирующей ткани - это активация защиты от возможности канцерогенеза ( ), что, в частности, связано с активацией апоптоза ( ) в результате активации синтеза p53. Кроме того окись азота регулирует работу сократительного белка миозина (), который, как известно, может играть ключевую роль в организации шаблона бластемы так как контролирует адгезию, поляризацию и миграцию клеток ( ). Эти функции миозин выполняет вслед за актином – мышечным белком, который выполняет роль инициатора этих процессов, обладая способностью к направленной полимеризации регулируемой Rho GTPазой Rho GTPases ( ). В связи с тем, что механизм защиты регенерирующей ткани от канцерогенеза важнейшее условие удачной регенерации, остановимся на нем немного подробнее. Организм, у которого активно обновляются ткани, рискует заболеть из-за чрезмерной пролиферации его клеток. Худшим вариантом такого заболевания является рак. Этому риску противостоят механизмы подавления опухолевого роста. Одним из таких механизмов является ускоренное старение дефектных клеток. В этом механизме задействованы: сигнальный путь регулируемый белком p53 ( ) и сигнальный путь регулируемый белками pRb и p16 Ink4a ( , ). Мутации в этих белках значительно повышают риск развития рака ( ). Вместе с тем именно эти белки играют важнейшую роль в ограничении способности клетки к неограниченному делению ( , ) и перепрограммированию в ИПСК ( , ). В экспериментах с генами Rb (ген подавляющий опухоль - ретинобластому) и Ink4A (продуктом гена ARF, который присутствует в геноме высших позвоночных животных, но отсутствует у тритона и саламандры) было показано, что инактивация Rb приводит к активации деления мышечных клеток у саламандры (но не у млекопитающих). Совместное отключение (с помощью «нокдауна генов» интерференционной РНК) и ARF и Rb приводит к дедифференцировке клеток млекопитающего (мыши) и их активной пролиферации как у раковых клеток. По мнению авторов исследования, за появление в геноме млекопитающих гена ARF они заплатили потерей способности к такой регенерации, как у тритона и саламандры ( ). Важно отметить, что в этом исследовании удалось превратить вырезанный лазером миоцит, который уже потерял способность к делению, в миобласт, способный образовывать колонии миобластов, пригодных для дифференцировки in vivo в миофибриллы после трансплантации в мышцу. Если удастся показать, что такие выращенные в пробирке миобласты не приводят после трансплантации в организм к образованию в последующем опухоли и что они работоспособны и правильно интегрированы в мышцу, можно будет использовать этот метод в хирургической практике. Есть надежды, что можно будет лечить повреждения, вызванные огнестрельными ранениями мышц, или даже инфарктом миокарда, путем стимуляции образования прогениторных мышечных клеток с помощью инъекций интерференционных РНК ингибирующих Rb и ARF.
Часть. Стволовые клетки
правитьЧасть. Основы эпигенетики развития
правитьЦикл изменений метилирования ДНК в процессе развития
правитьПаттерн (характерное распределение, узор) метилирования ДНК возникает на ДНК под влиянием метилтрансфераз DNMT3A и DNMT3B, которые переносят метильную группу с S-аденозил-метионина на цитозин ДНК с образованием 5-метилцитозина (1), а затем сохраняется и поддерживается при клеточных делениях метилтрансферазой DNMT1 совместно с UHRF1 - ДНК-связывающим белком, распознающим 5-метилцитозин. Нарушение взаимодействия DNMT1 с UHRF1 малой молекулой NSC232003 приводит к общему деметилированию цитозинов на ДНК после деления клетки (2).

Рис. 1. После репликации двухспиральная синтезированная ДНК имеет метильные группы только на одной цепи. Чтобы скопировать паттерн метилированных цитозинов UHRF1 связывается с метилированной ДНК и метит гистон Н3 убиквитином (Ubi). Затем эту метку и белок UHRF1 распознает метилтрансфераза DNMT1 и сажает метильную группу на цитозин противоположной цепи.(3)
Важную роль в регуляции процессов метилирования ДНК по-видимому играет семейство микроРНК miR-29. При истощении miR-29a наблюдается общее понижение уровня метилирования ДНК. Следует отметить, что факторы способствующие старению и канцерогенезу, такие как c-Myc ( ), сигнальный путь хеджехог (hedgehog) и воспалительные процессы (NF-κB сигналы) ( ) подавляют синтез miR-29 ( ). В то же время блокировка miR-29a приводит к повышению эффективности перепрограммирования соматических клеток в индуцированные плюрипотентные стволовые клетки iPSC ( ), повышая пластичность клетки – её способность к трансформации (в том числе и в раковые клетки). Предполагается, что miR-29a является регулятором, который ингибирует как ДНК метилтрансферазы, так и деметилазы ( ). Так, например, принудительное повышение синтеза miR-29 подавляя активность ДНК метилтрансфераз DNMT3A и -B восстанавливало паттерн метилирования ДНК и таким образом индуцировало реэкспрессию заингибированных метилированием генов супрессоров опухоли таких как FHIT и WWOX, что приводило к ингибированию онкогенеза как in vitro, так и in vivo ( ). Важно отметить, что уровень miR-29 в старых клетках намного выше чем в молодых ( ) и что повышение активности miR-29 приводит понижению активности множества генов, способствующих росту тела, тем самым способствуя физиологическому замедлению и в конечном итоге прекращения роста тела ( ). Ингибирование miR-29 in vivo ускоряет экспрессию фенотипов старения ( ), в частности, процессы усиленного отложение коллагена и фиброза ( )
Ниже приведен рисунок, на котором в схематическом виде представлены известные на сегодняшний день ключевые ферменты, участвующие в механизмах «записи» и «стирания» генетической информации.

Рис 2. По (17). Упрощённая схема и участники механизмов «записи» и «стирания» эпигенетической информации. TrxG (Trithorax group) – комплекс белков группы Триторакс; PRC2 (Polycomb Repressive Complex 2) – репрессорный комплекс белков группы Поликомб; (Kdm3a, Kdm3b, Kdm4a, Kdm4b, Kdm5b, Kdm6a) – деметиллазы лизина гистонов; Hdac (Histone deacetylase of NuRD complex) – деацетиллазы гистонов; Ehmt2 (Euchromatic histone-lysine N-methyltransferase 2), Setdb1, Suv39h1/2 – метилтрансферазы лизина 9 в гистоне H3; TET1/2/3 (Ten-eleven translocation methylcytosine dioxygenases 1/2/3) – ферменты катализирующие окисление оснований 5-метилцитозина (5-mC) в ДНК до 5-гидроксиметилцитозина (5-hmC), что ведет к активному деметилированию этого цитозина в составе ДНК; Dmt1/3a/3b (Dnmt1, Dnmt3a, Dnmt3b) - метилтрансферазы ДНК. Из рисунка наглядно видно, что для эухроматина – хроматина на котором активно идет транскрипция РНК, характерны метки активного хроматина такие как ацетилированные гистоны, гистон H3K4me3 и ДНК с большей частью неметилированных цитозинов. Другая картина наблюдается на гетерохроматине – конденсированном хроматине, где синтез РНК заблокирован. На гетерохроматине преобладают метки не активного хроматина, такие как H3K9me3 и H3K27me3, а цитозины ДНК обычно метилированы.
Как меняется метилирование ДНК на протяжении жизни организма. Динамику этого процесса можно проследить по приведенным ниже двум рисункам и прочитав пояснительный текст к этим рисункам.

Рис 3. По (18). Эпигенетические модификации подвергаются перепрограммированию в течение жизненного цикла в два этапа: во время гаметогенеза (образования сперматозоидов у мужчин и ооцитов у женщин) и во время предимплантационного развития, когда оплодотворение дает сигнал для второго тура перепрограммирования. Отцовский геном подвергается активному деметилированию (см. следующий рисунок). В процессе первых делений генома зародыша пассивно деметилируется большая часть его ДНК (при этом наблюдается геномный импритинг – экспрессия и метилирование ДНК определённых генов поддерживается в зависимости от того, от отца или матери поступил ген). Такое пассивное деметилирование наблюдается вплоть до процесса образования бластоцисты. Метилирование de novo (заново) примерно совпадает с дифференциацией клеток на первые две линии клеток бластоцисты: трофобласты дающие начало плаценте и клетки внутренней клеточной массы, ДНК которых гиперметилирована в отличие от трофобластов. Эта ассиметрия в уровне метилирования ДНК сопровождается и ассиметрией в характере модификаций гистонов.

Рис 4. По (19) Из графика видно, что деметилирование сначала происходит на отцовской ДНК, а уже потом и на материнской. Последующая специализация клеток связана с её эпигенетическим закреплением в виде de novo метилирования ДНК.
ДНК в сперматозоидах и в ооцитах имеет высокую степень метилирования. Но на самых ранних этапах развития эмбриона, при образовании в результате оплодотворения зиготы – клетки, с которой начинается процесс развития нового организма, уровень метилирования ДНК снижается. Уже вскоре после оплодотворения отцовская ДНК в зиготе почти полностью деметилирована. Это деметилирование отцовской ДНК является ферментативным процессом, так как происходит в отсутствие репликации ДНК. Вслед за этим, но уже пассивно, в ходе клеточных делений, деметилируется и материнская ДНК. Небольшая часть ДНК в ходе этих процессов остаётся метилированной. К тому времени, когда эмбрион достигает стадии морулы, его геном почти полностью лишен метилирования ДНК ( ). Такое первоначальное снижение уровня метилирования ДНК, очевидно, предусмотрено природой для того, чтобы начать развитие организма с «чистого листа». В ходе последующего развития эмбриона начинается специализация клеток с её эпигенетическим закреплением в виде de novo (заново) метилирования ДНК ( ). Таким образом, «запись» об изменениях связанных с дифференцировкой закрепляется в эпигеноме клеток потомков зиготы ( ). При этом из одной оплодотворенной яйцеклетки человека в течение жизни образуется порядка 37000 миллиардов (37 триллиардов) различных клеток, из которых состоят 210 гистологически различимых тканей формирующих высокоспециализированные органы нашего организма ( ). Уже в ДНК эмбриональных стволовых клеток человека метилирование достигает максимума - метилировано почти 80% CpG, а оставшиеся неметилированными CpG остатки обычно расположены в островках CpGi, локализованных на генных промоторах ( ). Важность метилирования ДНК в процессе развития была подкреплена исследованиями, показавшими, что мыши, лишенные какой-либо из метилтрансфераз ДНК (DNMT1, Dnmt3a и DNMT3b) не являются жизнеспособными и умирают ( ). Эти и некоторые другие наблюдения, закрепили представление о том, что ключевые роли в процессах клеточной дифференцировки исполняют ферменты, влияющие на эпигенетические «записи» в виде метилирования ДНК и ферментативных модификаций гистонов.
Примечания
править- Okano M, Bell DW, Haber DA, Li E (1999). DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell; 99(3), 247-57.
- Myrianthopoulos, V., Cartron, P. F., Liutkevičiūtė, Z., Klimašauskas, S., Matulis, D., Bronner, C., ... & Mikros, E. (2016). Tandem virtual screening targeting the SRA domain of UHRF1 identifies a novel chemical tool modulating DNA methylation. European journal of medicinal chemistry, 114, 390-396. doi:10.1016/j.ejmech.2016.02.043 http://www.sciencedirect.com/science/article/pii/S0223523416301337
- Nishiyama, A., Yamaguchi, L., & Nakanishi, M. (2016). Regulation of maintenance DNA methylation via histone ubiquitylation. Journal of biochemistry, 159(1), 9-15. doi: 10.1093/jb/mvv113 http://jb.oxfordjournals.org/content/159/1/9.full
- Hofmann, J. W., Zhao, X., De Cecco, M., Peterson, A. L., Pagliaroli, L., Manivannan, J., ... & Li, X. (2015). Reduced expression of MYC increases longevity and enhances healthspan. Cell, 160(3), 477-488. doi:10.1016/j.cell.2014.12.016 http://www.sciencedirect.com/science/article/pii/S0092867414015864
- Saha, M. N., Abdi, J., Yang, Y., & Chang, H. (2016). miRNA-29a as a tumor suppressor mediates PRIMA-1Met-induced anti-myeloma activity by targeting c-Myc. Oncotarget, 7(6), 7149.
- Li, M., & Guan, H. (2016). Noncoding RNAs Regulating NF-κB Signaling. In The Long and Short Non-coding RNAs in Cancer Biology (pp. 317-336). Springer Singapore. DOI: 10.1007/978-981-10-1498-7_12 http://link.springer.com/chapter/10.1007/978-981-10-1498-7_12
- Mott, J. L., Kurita, S., Cazanave, S. C., Bronk, S. F., Werneburg, N. W., & Fernandez-Zapico, M. E. (2010). Transcriptional suppression of mir-29b-1/mir-29a promoter by c-Myc, hedgehog, and NF-kappaB. Journal of cellular biochemistry, 110(5), 1155-1164. DOI: 10.1002/jcb.22630 http://onlinelibrary.wiley.com/doi/10.1002/jcb.22630/full
- Hysolli, E., Tanaka, Y., Su, J., Kim, K. Y., Zhong, T., Janknecht, R., ... & Jung, Y. W. (2016). Regulation of the DNA Methylation Landscape in Human Somatic Cell Reprogramming by the miR-29 Family. Stem Cell Reports. doi:10.1016/j.stemcr.2016.05.014 http://www.sciencedirect.com/science/article/pii/S2213671116300698
- Fráguas, M. S., Eggenschwiler, R., Hoepfner, J., dos Santos Schiavinato, J. L., Haddad, R., Oliveira, L. H. B., ... & Cantz, T. (2017). MicroRNA-29 impairs the early phase of reprogramming process by targeting active DNA demethylation enzymes and Wnt signaling. Stem Cell Research, 19, 21-30. https://doi.org/10.1016/j.scr.2016.12.020
- Jia, L. F., Zheng, Y. F., Lyu, M. Y., Huang, Y. P., & Gan, Y. H. (2016). miR-29b upregulates miR-195 by targeting DNMT3B in tongue squamous cell carcinoma.Science Bulletin, 61(3), 212-219.
- Fabbri, M., Garzon, R., Cimmino, A., Liu, Z., Zanesi, N., Callegari, E., ... & Volinia, S. (2007). MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proceedings of the National Academy of Sciences, 104(40), 15805-15810. doi: 10.1073/pnas.0707628104
- Martinez, I., Cazalla, D., Almstead, L. L., Steitz, J. A., & DiMaio, D. (2011). miR-29 and miR-30 regulate B-Myb expression during cellular senescence.Proceedings of the National Academy of Sciences, 108(2), 522-527. • doi: 10.1073/pnas.1017346108 http://www.pnas.org/content/108/2/522.short
- Li, T., Yan, X., Jiang, M., & Xiang, L. (2016). The comparison of microRNA profile of the dermis between the young and elderly. Journal of dermatological science,82(2), 75-83. doi:10.1016/j.jdermsci.2016.01.005 http://www.sciencedirect.com/science/article/pii/S0923181116300068
- Kamran, F., Andrade, A. C., Nella, A. A., Clokie, S. J., Rezvani, G., Nilsson, O., ... & Lui, J. C. (2015). Evidence That Up-Regulation of MicroRNA-29 Contributes to Postnatal Body Growth Deceleration. Molecular Endocrinology, 29(6), 921-932. .doi.:10.1210/me.2015-1047 http://press.endocrine.org/doi/10.1210/me.2015-1047
- Cellerino, A. (2017). From the bush to the bench: A short-lived african fish reveals novel aspects of the genetic control of development and aging. Mechanisms of Development, 145, S10.
- Wang, B., Komers, R., Carew, R., Winbanks, C. E., Xu, B., Herman-Edelstein, M., ... & Cooper, M. E. (2012). Suppression of microRNA-29 expression by TGF-β1 promotes collagen expression and renal fibrosis. Journal of the American Society of Nephrology, 23(2), 252-265.
- Gładych, M., Andrzejewska, A., Oleksiewicz, U., & Estécio, M. R. (2015). Epigenetic mechanisms of induced pluripotency. Contemp Oncol (Pozn), 19(1A), A30-A38. DOI: 10.5114/wo.2014.47135
- Morgan, H. D., Santos, F., Green, K., Dean, W., & Reik, W. (2005). Epigenetic reprogramming in mammals. Human molecular genetics, 14(suppl 1), R47-R58.
- Marcho, C., Cui, W., & Mager, J. (2015). Epigenetic dynamics during preimplantation development. Reproduction, 150(3), R109-R120.
- Santos F, Hendrich B, Reik W, Dean W (2002). Dynamic reprogramming of DNA methylation in the early mouse embryo. Dev Biol; 241(1), 172-182.
- Lim, C. Y., Knowles, B. B., Solter, D., & Messerschmidt, D. M. (2016). Epigenetic Control of Early Mouse Development. Current Topics in Developmental Biology. doi:10.1016/bs.ctdb.2016.05.002 http://www.sciencedirect.com/science/article/pii/S0070215316301235
- Amouroux, R., Nashun, B., Shirane, K., Nakagawa, S., Hill, P. W., D’Souza, Z., ... & Encheva, V. (2016). De novo DNA methylation drives 5hmC accumulation in mouse zygotes. Nature cell biology, 18(2), 225-233. doi:10.1038/ncb3296 http://www.nature.com/ncb/journal/v18/n2/abs/ncb3296.html
- Bianconi E, Piovesan A,Facchin F (2013). An estimation of the number of cells in the human body. Ann. Hum. Biol. 40,463–471
- Chen PY, Feng S, Joo JW, Jacobsen SE, Pellegrini M (2011). A comparative analysis of DNA methylation across human embryonic stem cell lines. Genome Biol; 12(7), R62
- Li E, Bestor TH, Jaenisch R. (1992). Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell; 69: 915-26.
- Okano M, Bell DW, Haber DA, Li E.(1999). DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell; 99: 247-57